Genetics and Human Behaviour
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cognitive Ability and Education: How Behavioural Genetic Research Has Advanced Our Knowledge and Understanding of Their Association
Cognitive ability and education: how behavioural genetic research has advanced our knowledge and understanding of their association Margherita Malanchini1,2,3, Kaili Rimfeld2, Andrea G. Allegrini2, Stuart J. Ritchie2, & Robert Plomin2 1 Department of Biological and Experimental Psychology, Queen Mary University of London, United Kingdom. 2 Social, Genetic and Developmental Psychiatry Centre, Institute of Psychiatry, Psychology and Neuroscience, King’s College London, United Kingdom. 3 Population Research Center, The University of Texas at Austin, United States. Correspondence: Margherita Malanchini, Department of Biological and Experimental Psychology, Queen Mary University of London, Office 2.02, G.E. Fogg Building, Mile End Road, London E1 4NS. E-mail: [email protected] Abstract Cognitive ability and educational success predict positive outcomes across the lifespan, from higher earnings to better health and longevity. The shared positive outcomes associated with cognitive ability and education are emblematic of the strong interconnections between them. Part of the observed associations between cognitive ability and education, as well as their links with wealth, morbidity and mortality, are rooted in genetic variation. The current review evaluates the contribution of decades of behavioural genetic research to our knowledge and understanding of the biological and environmental basis of the association between cognitive ability and education. The evidence reviewed points to a strong genetic basis in their association, observed from middle childhood to old age, which is amplified by environmental experiences. In addition, the strong stability and heritability of educational success are not driven entirely by cognitive ability. This highlights the contribution of other educationally relevant noncognitive characteristics. Considering both cognitive and noncognitive skills as well as their biological and environmental underpinnings will be fundamental in moving towards a comprehensive, evidence-based model of education. -

AIH Chapter 2: Human Behavior
Aviation Instructor's Handbook (FAA-H-8083-9) Chapter 2: Human Behavior Introduction Derek’s learner, Jason, is very smart and able to retain a lot of information, but has a tendency to rush through the less exciting material and shows interest and attentiveness only when performing tasks that he finds to be interesting. This concerns Derek because he is worried that Jason will overlook many important details and rush through procedures. For a homework assignment Jason was told to take a very thorough look at Preflight Procedures and that for his next flight lesson they would discuss each step in detail. As Derek predicted, Jason found this assignment to be boring and was not prepared. Derek knows that Jason is a “thrill seeker” as he talks about his business, which is a wilderness adventure company. Derek wants to find a way to keep Jason focused and help him find excitement in all areas of learning so that he will understand the complex art of flying and aircraft safety. Learning is the acquisition of knowledge or understanding of a subject or skill through education, experience, practice, or study. This chapter discusses behavior and how it affects the learning process. An instructor seeks to understand why people act the way they do and how people learn. An effective instructor uses knowledge of human behavior, basic human needs, the defense mechanisms humans use that prevent learning, and how adults learn in order to organize and conduct productive learning activities. Definitions of Human Behavior The study of human behavior is an attempt to explain how and why humans function the way they do. -

Vide O Clipe: Forças E Sensações No Caos
UNIVERSIDADE ESTADUAL DE CAMPINAS FACULDADE DE EDUCAÇÃO DISSERTAÇÃO DE MESTRADO Vide o clipe: forças e sensações no caos Autor: Pamela Zacharias Sanches Oda Orientador: Prof. Dr. Antonio Carlos Rodrigues de Amorim Este exemplar corresponde à redação final da Dissertação defendida por Pamela Zacharias Sanches Oda e aprovada pela Comissão Julgadora. Data: 15 / 02 / 2011 2011 ii iii Ao Lucas, Meu amor, amigo e companheiro, que possibilitou através de seu incentivo, de seu olhar crítico e de sua imensa paciência a concretização deste trabalho. iv AGRADECIMENTOS Agradecer é estar em estado de graça. Assim me sinto ao traçar as últimas linhas dessa dissertação. Nesse momento, rememoro todo o trajeto do mestrado e percebo que o que me trouxe aqui se iniciou bem antes da escrita do projeto, bem antes de fazer minha inscrição para o processo seletivo. O início de tudo está em um tempo não situável e deve-se a tantos corpos que não haveria espaço para agradecê-los nestas páginas. Por isso o estado de graça, a gratidão ampla e imensurável por estar onde estou e poder deixar um registro de um pedacinho de minha vida no mundo, mesmo que perdido entre as estantes de uma biblioteca acadêmica. Isso porque este texto é apenas a materialidade simbólica de tudo o que este trabalho me trouxe e vai além, muito além do que ele apresenta. Agradeço, portanto, ao prof. dr. Antônio Carlos de Amorim, por ter se interessado em me orientar e ter me possibilitado tantos encontros teóricos e principalmente humanos; ao grupo de pesquisa OLHO e, em especial ,ao Humor -

To What Extent Does the Heritability Of
1 Bonamy R. Oliver & Alison Pike University of Sussex 'To what extent does the heritability of known features of development such as intelligence limit the potential of policy that attempts to lessen intergenerational inequalities such as early intervention, social mobility, child poverty or the social class attainment gap'. In the current report, we argue that the extent to which aspects of human development are heritable has little impact on the importance of policy that aims to ameliorate the adverse effects of environmental or intergenerational inequalities. We first address common misconceptions about heritability and behavioural genetics that can lead to inappropriate assertions about research findings and their implications for policy. We then move to demonstrate our vision of what behavioural genetics can offer early intervention science and attempts to lessen intergenerational inequality. What is behavioural genetics? Behavioural genetic research uses a variety of quasi-experimental designs to understand the role of genetic and environmental influence on variation in psychological traits such as general intelligence (e.g., Deary, Johnson, & Houlihan, 2009), disruptive behaviour (e.g., Rhee, & Waldman, 2002), and well-being (e.g., Bartels, 2015). The extent to which variance in a given trait is explained by genetic variance is termed ‘heritability’, and is explained further below. Shared environmental influences refer to experiences that contribute to sibling similarity, while non- shared environmental influences refer to differences in experience that contribute to differences between siblings in a family. The most common of these research designs are twin and adoption 2 studies. The twin design compares monozygotic (MZ: ‘identical’) and dizygotic (DZ: fraternal, ‘not identical’) twin similarity for specific psychological traits. -

The Psychology of Music Haverford College Psychology 303
The Psychology of Music Haverford College Psychology 303 Instructor: Marilyn Boltz Office: Sharpless 407 Contact Info: 610-896-1235 or [email protected] Office Hours: before class and by appointment Course Description Music is a human universal that has been found throughout history and across different cultures of the world. Why, then, is music so ubiquitous and what functions does it serve? The intent of this course is to examine this question from multiple psychological perspectives. Within a biological framework, it is useful to consider the evolutionary origins of music, its neural substrates, and the development of music processing. The field of cognitive psychology raises questions concerning the relationship between music and language, and music’s ability to communicate emotive meaning that may influence visual processing and body movement. From the perspectives of social and personality psychology, music can be argued to serve a number of social functions that, on a more individual level, contribute to a sense of self and identity. Lastly, musical behavior will be considered in a number of applied contexts that include consumer behavior, music therapy, and the medical environment. Prerequisites: Psychology 100, 200, and at least one advanced 200-level course. Biological Perspectives A. Evolutionary Origins of Music When did music evolve in the overall evolutionary scheme of events and why? Does music serve any adaptive purposes or is it, as some have argued, merely “auditory cheesecake”? What types of evidence allows us to make inferences about the origins of music? Reading: Thompson, W.F. (2009). Origins of Music. In W.F. Thompson, Music, thought, and feeling: Understanding the psychology of music. -

Social Psychology Glossary
Social Psychology Glossary This glossary defines many of the key terms used in class lectures and assigned readings. A Altruism—A motive to increase another's welfare without conscious regard for one's own self-interest. Availability Heuristic—A cognitive rule, or mental shortcut, in which we judge how likely something is by how easy it is to think of cases. Attractiveness—Having qualities that appeal to an audience. An appealing communicator (often someone similar to the audience) is most persuasive on matters of subjective preference. Attribution Theory—A theory about how people explain the causes of behavior—for example, by attributing it either to "internal" dispositions (e.g., enduring traits, motives, values, and attitudes) or to "external" situations. Automatic Processing—"Implicit" thinking that tends to be effortless, habitual, and done without awareness. B Behavioral Confirmation—A type of self-fulfilling prophecy in which people's social expectations lead them to behave in ways that cause others to confirm their expectations. Belief Perseverance—Persistence of a belief even when the original basis for it has been discredited. Bystander Effect—The tendency for people to be less likely to help someone in need when other people are present than when they are the only person there. Also known as bystander inhibition. C Catharsis—Emotional release. The catharsis theory of aggression is that people's aggressive drive is reduced when they "release" aggressive energy, either by acting aggressively or by fantasizing about aggression. Central Route to Persuasion—Occurs when people are convinced on the basis of facts, statistics, logic, and other types of evidence that support a particular position. -

Vrije Universiteit Amsterdam Objects and Their Stories 1985-1990 Nick and Carst and the Netherlands Twin Register
Vrije Universiteit Amsterdam objects and their stories 1985-1990 Nick and Carst and the Netherlands Twin Register There are many research institutes at VU Amsterdam. These institutes often bring together various disciplines around a specific theme. One of the more unusual topics is research on twins, which has been carried out at VU Amsterdam since the 1980s. This institute is represented in the collections in a unique way: with a work of art. Art curator Wende Wallert and Professor Dorret Boomsma from the Twin Institute tell us more. Nick and Carst By Wende Wallert In 2016, photographers Monique Eller and Bodine Koopmans made a series of portraits about the twin phenomenon entitled ‘DOUBLE DUTCH’, in collaboration with the NTR and VU Amsterdam. The results of their work were displayed in an exhibition in the Main Building. Set against a muted background, the portrait of Nick and Carst beautifully reveals their similarities and differences as well as the special bond that twins share. Monique Eller and Bodine Koopmans, Nick and Carst, 2016, VU VU Amsterdam aims to stimulate the interaction Amsterdam Art Collection between art and science. Like science, art offers us new insights, but from the imagination. In its own gallery space called WONDER, VU Amsterdam organises exhibitions and events to share current artistic and scientific research with the public. The Netherlands Twin Register By Dorret Boomsma I began my PhD studies at Vrije Universiteit Amsterdam in 1983, after completing a doctoral programme there and obtaining an MA at the Institute for Behavioural Genetics in Boulder, Colorado. It was in Boulder that I became aware of the value of twin studies. -

Mediated Music Makers. Constructing Author Images in Popular Music
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Helsingin yliopiston digitaalinen arkisto Laura Ahonen Mediated music makers Constructing author images in popular music Academic dissertation to be publicly discussed, by due permission of the Faculty of Arts at the University of Helsinki in auditorium XII, on the 10th of November, 2007 at 10 o’clock. Laura Ahonen Mediated music makers Constructing author images in popular music Finnish Society for Ethnomusicology Publ. 16. © Laura Ahonen Layout: Tiina Kaarela, Federation of Finnish Learned Societies ISBN 978-952-99945-0-2 (paperback) ISBN 978-952-10-4117-4 (PDF) Finnish Society for Ethnomusicology Publ. 16. ISSN 0785-2746. Contents Acknowledgements. 9 INTRODUCTION – UNRAVELLING MUSICAL AUTHORSHIP. 11 Background – On authorship in popular music. 13 Underlying themes and leading ideas – The author and the work. 15 Theoretical framework – Constructing the image. 17 Specifying the image types – Presented, mediated, compiled. 18 Research material – Media texts and online sources . 22 Methodology – Social constructions and discursive readings. 24 Context and focus – Defining the object of study. 26 Research questions, aims and execution – On the work at hand. 28 I STARRING THE AUTHOR – IN THE SPOTLIGHT AND UNDERGROUND . 31 1. The author effect – Tracking down the source. .32 The author as the point of origin. 32 Authoring identities and celebrity signs. 33 Tracing back the Romantic impact . 35 Leading the way – The case of Björk . 37 Media texts and present-day myths. .39 Pieces of stardom. .40 Single authors with distinct features . 42 Between nature and technology . 45 The taskmaster and her crew. -

Holmes CV Current.Pdf
Biosketch NAME POSITION TITLE Andrew Holmes, Ph.D. Acting Chief, Section on Behavioral Science and Genetics, Laboratory for Integrative Neuroscience, National Institute on Alcohol Abuse and Alcoholism, National Institutes of Health EDUCATION/TRAINING DEGREE INSTITUTION AND LOCATION (if applicable) YEAR(s) FIELD OF STUDY Department of Psychology, University of B.A. (Hons) 1992-1995 Psychology Newcastle upon Tyne, UK Department of Psychology, University of Ph.D. 1995-1999 Behavioral Pharmacology Leeds, UK (Mentor, RJ Rodgers) National Institute of Mental Health, Visiting Fellow 1999-2003 Behavioral Neuroscience National Institutes of Health, USA (Mentor, JN Crawley) Awards 2008: United States - Israel Binational Science Foundation Four-Year Grant Award 2008: Alcoholism and Stress: A Framework for Future Treatment Strategies, Young Investigator Award 2007: Senior Preclinical Wyeth Psychopharmacology Award, British Association of Psychopharmacology 2006: Young Scientist Award, International Behavioural and Neural Genetics Society 2005: National Alliance for Research on Schizophrenia and Depression Young Investigator Award 2004: NIH Pre-IRTA Committee Awards for outstanding contributions to the post-baccalaureate training and education 2003: Anxiety Disorders Association of America/American College of Neuropsychopharmacology Career Development Travel Award 2003: Howard Hughes Medical Institute Student Mentor Award 2002: NIH Fellows Award for Research Excellence 2002: New York Academy of Sciences Travel Award 2001: American College of Neuropsychopharmacology/Bristol-Myers -

ONFERENCE PROG the 13Th International
ONFERENCE PROG C The 13th International Congress on Twin Studies June 4-7, 2010, Seoul, Korea cv mu"' f6ARtt Seoul c.cio4"J' RA7°J2-- U/1 TOURISM ORGANIZATION FRIDAY (JUNE 4) 18:30 -21:00 ICTS Welcome Reception (Regency Room) SATURDAY (JUNE 5) 7:30 - 8:15 Breakfast (Regency Room) Symposium : Genetics of high cognitive abilities Regency Room 815 - 9:45 Organizer: Robert Plomin Robert Plomin & Claire M. A. Haworth 8:15 Genetics of high cognitive abilities : An overview John Hewitt, Angela Brant, Robin Corley, Sally Wadsworth & John DeFries 8:30 A closer look at the developmental etiology of high IQ Matt McGue, Robert M. Kirkpatrick, Michael Miller, & William G. Iacono 8:45 The University of Minnesota Initiative on the Genetics of High Cognitive Ability Allan McRae, Margie Wright, Narelle Hansell, Peter Visscher, Grant Montgomery, & Nicholas G. Martin 9:00 Is high cognitive ability associated with a lower genomic burden of copy number variants? Dorret I. Boomsma, Catherina E.M. van Beijsterveldt, Inge van Soelen , Sanja Franio, Conor V. Dolan, Denny Borsboom,& M. Bartels 9:15 Genetic analysis of longitudinally measured IQ, educational attainment and educational level in Dutch twin-sib samples Paper Session : Twin studies of personality & psychological wellbeing Namsan Ill 8:15 - 9:45 Chair: Corrado Fagnani Christian Kandler, Wiebke Bleidorn, & Rainer Riemann 8:15 Sources of cumulative continuity in personality: A longitudinal multiple-rater twin study Veselka, L., Schermer, J.A., Martin, R. A., & Vernon, P.A. 8:30 The heritability of general factor of personality extracted in four different datasets Antonella Gigantesco, Corrado Fagnani, Guido Alessandri, Emanuela Medda, 8:45 Emanuele Tarolla, & Maria Antonietta Stazi An Italian twin study on psychological well-being in young adulthood Meike Bartels, Gonneke, A.H.M. -
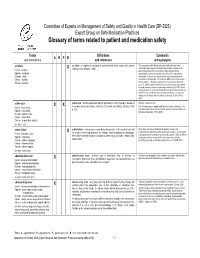
Glossary of Terms Related to Patient and Medication Safety
Committee of Experts on Management of Safety and Quality in Health Care (SP-SQS) Expert Group on Safe Medication Practices Glossary of terms related to patient and medication safety Terms Definitions Comments A R P B and translations and references and synonyms accident accident : an unplanned, unexpected, and undesired event, usually with adverse “For many years safety officials and public health authorities have Xconsequences (Senders, 1994). discouraged use of the word "accident" when it refers to injuries or the French : accident events that produce them. An accident is often understood to be Spanish : accidente unpredictable -a chance occurrence or an "act of God"- and therefore German : Unfall unavoidable. However, most injuries and their precipitating events are Italiano : incidente predictable and preventable. That is why the BMJ has decided to ban the Slovene : nesreča word accident. (…) Purging a common term from our lexicon will not be easy. "Accident" remains entrenched in lay and medical discourse and will no doubt continue to appear in manuscripts submitted to the BMJ. We are asking our editors to be vigilant in detecting and rejecting inappropriate use of the "A" word, and we trust that our readers will keep us on our toes by alerting us to instances when "accidents" slip through.” (Davis & Pless, 2001) active error X X active error : an error associated with the performance of the ‘front-line’ operator of Synonym : sharp-end error French : erreur active a complex system and whose effects are felt almost immediately. (Reason, 1990, This definition has been slightly modified by the Institute of Medicine : “an p.173) error that occurs at the level of the frontline operator and whose effects are Spanish : error activo felt almost immediately.” (Kohn, 2000) German : aktiver Fehler Italiano : errore attivo Slovene : neposredna napaka see also : error active failure active failures : actions or processes during the provision of direct patient care that Since failure is a term not defined in the glossary, its use is not X recommended. -

Holding Back the Genes: Limitations of Research Into Canine Behavioural
van Rooy et al. Canine Genetics and Epidemiology 2014, 1:7 http://www.cgejournal.org/content/1/1/7 REVIEW Open Access Holding back the genes: limitations of research into canine behavioural genetics Diane van Rooy*, Elizabeth R Arnott, Jonathan B Early, Paul McGreevy and Claire M Wade Abstract Canine behaviours that are both desirable and undesirable to owners have a demonstrable genetic component. Some behaviours are breed-specific, such as the livestock guarding by maremmas and flank sucking seen in Dobermanns. While the identification of genes responsible for common canine diseases is rapidly advancing, those genes underlying behaviours remain elusive. The challenges of accurately defining and measuring behavioural phenotypes remain an obstacle, and the use of variable phenotyping methods has prevented meta-analysis of behavioural studies. International standardised testing protocols and terminology in canine behavioural evaluations should facilitate selection against behavioural disorders in the modern dog and optimise breeding success and performance in working dogs. This review examines the common hurdles faced by researchers of behavioural genetics and the current state of knowledge. Keywords: Behaviour, Dog, Genetics, Heritability Lay summary against behavioural disorders in the dog, and optimise All canine behaviour, whether desirable or undesirable breeding success and performance in working dogs. to owners, has a genetic component. Studies of “showing This review examines the common hurdles faced by eye” and “bark” which compared the behaviour of off- researchers of behavioural genetics and the current state spring with parents concluded that behaviour does not of knowledge. follow simple Mendelian inheritance. Some behaviours are breed-specific, such as “livestock guarding” by mar- Introduction emmas and “flank sucking” seen in Dobermanns.