Association of Dopaminergic/Gabaergic Genes with Attention Deficit Hyperactivity Disorder in Children
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

1 Evidence for Gliadin Antibodies As Causative Agents in Schizophrenia
1 Evidence for gliadin antibodies as causative agents in schizophrenia. C.J.Carter PolygenicPathways, 20 Upper Maze Hill, Saint-Leonard’s on Sea, East Sussex, TN37 0LG [email protected] Tel: 0044 (0)1424 422201 I have no fax Abstract Antibodies to gliadin, a component of gluten, have frequently been reported in schizophrenia patients, and in some cases remission has been noted following the instigation of a gluten free diet. Gliadin is a highly immunogenic protein, and B cell epitopes along its entire immunogenic length are homologous to the products of numerous proteins relevant to schizophrenia (p = 0.012 to 3e-25). These include members of the DISC1 interactome, of glutamate, dopamine and neuregulin signalling networks, and of pathways involved in plasticity, dendritic growth or myelination. Antibodies to gliadin are likely to cross react with these key proteins, as has already been observed with synapsin 1 and calreticulin. Gliadin may thus be a causative agent in schizophrenia, under certain genetic and immunological conditions, producing its effects via antibody mediated knockdown of multiple proteins relevant to the disease process. Because of such homology, an autoimmune response may be sustained by the human antigens that resemble gliadin itself, a scenario supported by many reports of immune activation both in the brain and in lymphocytes in schizophrenia. Gluten free diets and removal of such antibodies may be of therapeutic benefit in certain cases of schizophrenia. 2 Introduction A number of studies from China, Norway, and the USA have reported the presence of gliadin antibodies in schizophrenia 1-5. Gliadin is a component of gluten, intolerance to which is implicated in coeliac disease 6. -
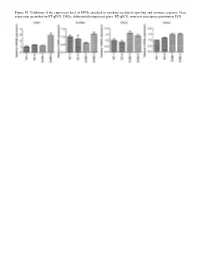
Figure S1. Validation of the Expression Level of Degs Enriched in Cytokine‑Mediated Signaling and Immune Response
Figure S1. Validation of the expression level of DEGs enriched in cytokine‑mediated signaling and immune response. Gene expression quantified by RT‑qPCR. DEGs, differentially expressed genes; RT‑qPCR, reverse‑transcription quantitative PCR. Figure S2. Validation of STAU1‑regulated AS events. IGV‑Sashimi plot revealed (A‑C) three A5SS AS events in three different genes. Reads distribution of each AS event was plotted in the left panel with the transcripts of each gene shown below. The sche‑ matic diagrams depict the structures of two AS events, AS1 (purple line) and AS2 (green line). The exon sequences are denoted by black boxes, the intron sequences by a horizontal line (right panel). RNA‑seq quantification and RT‑qPCR validation of ASEs are presented in the panels on the right. STAU1, double‑stranded RNA‑binding protein Staufen homolog 1; AS, alternative splicing; A5SS, alternative 5'splice site; RNA‑seq, RNA sequencing; RT‑qPCR, reverse‑transcription quantitative PCR. Error bars represent mean ± SEM. *P<0.05. Table SI. Primers used in gene validation experiments. IFIT2‑F CAGCCTACGGCAACTAAA IFIT2‑R GAGCCTTCTCAAAGCACA IFIT3‑F ACACCAAACAATGGCTAC IFIT3‑R TGGACAAACCCTCTAAAC OASL‑F AATGGTGACCGTGATGGG OASL‑R ACCTGAGGATGGAGCAGAG IFI27‑F TTCACTGCGGCGGGAATC IFI27‑R TGGCTGCTATGGAGGACGAG S1PR4‑F TGCTGAAGACGGTGCTGATG S1PR4‑R TGCGGAAGGAGTAGATGATGG CCL5‑F ACGACTGCTGGGTTGGAG CCL5‑R ACCCTGCTGCTTTGCCTA CCL2‑F CTAACCCAGAAACATCCAAT CCL2‑R GCTATGAGCAGCAGGCAC CD44‑F TGGAGGACAGAAAGCCAAGT CD44‑R TTCGCAATGAAACAATCAGTAG PLEKHG2‑M/As‑F CCAAAAGTAAGCCTGTCC PLEKHG2‑M‑R -

G Protein-Coupled Receptors
S.P.H. Alexander et al. The Concise Guide to PHARMACOLOGY 2015/16: G protein-coupled receptors. British Journal of Pharmacology (2015) 172, 5744–5869 THE CONCISE GUIDE TO PHARMACOLOGY 2015/16: G protein-coupled receptors Stephen PH Alexander1, Anthony P Davenport2, Eamonn Kelly3, Neil Marrion3, John A Peters4, Helen E Benson5, Elena Faccenda5, Adam J Pawson5, Joanna L Sharman5, Christopher Southan5, Jamie A Davies5 and CGTP Collaborators 1School of Biomedical Sciences, University of Nottingham Medical School, Nottingham, NG7 2UH, UK, 2Clinical Pharmacology Unit, University of Cambridge, Cambridge, CB2 0QQ, UK, 3School of Physiology and Pharmacology, University of Bristol, Bristol, BS8 1TD, UK, 4Neuroscience Division, Medical Education Institute, Ninewells Hospital and Medical School, University of Dundee, Dundee, DD1 9SY, UK, 5Centre for Integrative Physiology, University of Edinburgh, Edinburgh, EH8 9XD, UK Abstract The Concise Guide to PHARMACOLOGY 2015/16 provides concise overviews of the key properties of over 1750 human drug targets with their pharmacology, plus links to an open access knowledgebase of drug targets and their ligands (www.guidetopharmacology.org), which provides more detailed views of target and ligand properties. The full contents can be found at http://onlinelibrary.wiley.com/doi/ 10.1111/bph.13348/full. G protein-coupled receptors are one of the eight major pharmacological targets into which the Guide is divided, with the others being: ligand-gated ion channels, voltage-gated ion channels, other ion channels, nuclear hormone receptors, catalytic receptors, enzymes and transporters. These are presented with nomenclature guidance and summary information on the best available pharmacological tools, alongside key references and suggestions for further reading. -

Multi-Functionality of Proteins Involved in GPCR and G Protein Signaling: Making Sense of Structure–Function Continuum with In
Cellular and Molecular Life Sciences (2019) 76:4461–4492 https://doi.org/10.1007/s00018-019-03276-1 Cellular andMolecular Life Sciences REVIEW Multi‑functionality of proteins involved in GPCR and G protein signaling: making sense of structure–function continuum with intrinsic disorder‑based proteoforms Alexander V. Fonin1 · April L. Darling2 · Irina M. Kuznetsova1 · Konstantin K. Turoverov1,3 · Vladimir N. Uversky2,4 Received: 5 August 2019 / Revised: 5 August 2019 / Accepted: 12 August 2019 / Published online: 19 August 2019 © Springer Nature Switzerland AG 2019 Abstract GPCR–G protein signaling system recognizes a multitude of extracellular ligands and triggers a variety of intracellular signal- ing cascades in response. In humans, this system includes more than 800 various GPCRs and a large set of heterotrimeric G proteins. Complexity of this system goes far beyond a multitude of pair-wise ligand–GPCR and GPCR–G protein interactions. In fact, one GPCR can recognize more than one extracellular signal and interact with more than one G protein. Furthermore, one ligand can activate more than one GPCR, and multiple GPCRs can couple to the same G protein. This defnes an intricate multifunctionality of this important signaling system. Here, we show that the multifunctionality of GPCR–G protein system represents an illustrative example of the protein structure–function continuum, where structures of the involved proteins represent a complex mosaic of diferently folded regions (foldons, non-foldons, unfoldons, semi-foldons, and inducible foldons). The functionality of resulting highly dynamic conformational ensembles is fne-tuned by various post-translational modifcations and alternative splicing, and such ensembles can undergo dramatic changes at interaction with their specifc partners. -

Download (PDF)
Supplemental Information Biological and Pharmaceutical Bulletin Promoter Methylation Profiles between Human Lung Adenocarcinoma Multidrug Resistant A549/Cisplatin (A549/DDP) Cells and Its Progenitor A549 Cells Ruiling Guo, Guoming Wu, Haidong Li, Pin Qian, Juan Han, Feng Pan, Wenbi Li, Jin Li, and Fuyun Ji © 2013 The Pharmaceutical Society of Japan Table S1. Gene categories involved in biological functions with hypomethylated promoter identified by MeDIP-ChIP analysis in lung adenocarcinoma MDR A549/DDP cells compared with its progenitor A549 cells Different biological Genes functions transcription factor MYOD1, CDX2, HMX1, THRB, ARNT2, ZNF639, HOXD13, RORA, FOXO3, HOXD10, CITED1, GATA1, activity HOXC6, ZGPAT, HOXC8, ATOH1, FLI1, GATA5, HOXC4, HOXC5, PHTF1, RARB, MYST2, RARG, SIX3, FOXN1, ZHX3, HMG20A, SIX4, NR0B1, SIX6, TRERF1, DDIT3, ASCL1, MSX1, HIF1A, BAZ1B, MLLT10, FOXG1, DPRX, SHOX, ST18, CCRN4L, TFE3, ZNF131, SOX5, TFEB, MYEF2, VENTX, MYBL2, SOX8, ARNT, VDR, DBX2, FOXQ1, MEIS3, HOXA6, LHX2, NKX2-1, TFDP3, LHX6, EWSR1, KLF5, SMAD7, MAFB, SMAD5, NEUROG1, NR4A1, NEUROG3, GSC2, EN2, ESX1, SMAD1, KLF15, ZSCAN1, VAV1, GAS7, USF2, MSL3, SHOX2, DLX2, ZNF215, HOXB2, LASS3, HOXB5, ETS2, LASS2, DLX5, TCF12, BACH2, ZNF18, TBX21, E2F8, PRRX1, ZNF154, CTCF, PAX3, PRRX2, CBFA2T2, FEV, FOS, BARX1, PCGF2, SOX15, NFIL3, RBPJL, FOSL1, ALX1, EGR3, SOX14, FOXJ1, ZNF92, OTX1, ESR1, ZNF142, FOSB, MIXL1, PURA, ZFP37, ZBTB25, ZNF135, HOXC13, KCNH8, ZNF483, IRX4, ZNF367, NFIX, NFYB, ZBTB16, TCF7L1, HIC1, TSC22D1, TSC22D2, REXO4, POU3F2, MYOG, NFATC2, ENO1, -

GPCR Expression Profiles Were Determined Using
Supplemental Figures and Tables for Tischner et al., 2017 Supplemental Figure 1: GPCR expression profiles were determined using the NanoString nCounter System in 250 ng of pooled cell RNA obtained from freshly isolated CD4 T cells from naïve lymph nodes (CD4ln), spinal cord infiltrating CD4 T cells at peak EAE disease (CD4sc), and primary lung endothelial cells (luEC). Supplemental Figure 2: Array design and quality controls. A, Sorted leukocytes or endothelial cells were subjected to single‐cell expression analysis and re‐evaluated based on the expression of various identity‐defining genes. B, Expression of identity‐defining and quality control genes after deletion of contaminating or reference gene‐negative cells. Expression data are calculated as 2(Limit of detection(LoD) Ct – sample Ct) ; LoD Ct was set to 24. Supplemental Figure 3: Overview over GPCR expression frequencies in different freshly isolated immune cell populations and spinal cord endothelial cells as determined by single cell RT‐PCR. Abbreviations: CD4ln‐Tcon/CD4ln‐Treg, conventional (con) and regulatory (reg) CD4 T cells from lymph nodes (CD4ln) of naïve mice; CD4dr/CD4sc, CD4 T cells from draining lymph nodes (dr) or spinal cord (sc) at peak EAE disease; CD4spn2D/ CD4spn2DTh1/ CD4spn2DTh17, splenic CD4 T cells from 2D2 T cell receptor transgenic mice before (2D) and after in vitro differentiation towards Th1 (2DTh1) or Th17 (2DTh17); MonoSpn, splenic monocytes; CD11b_sc, spinal cord infiltrating CD11b‐ positive cells; sc_microglia, Ccr2neg,Cx3cr1pos microglia from spinal cord at peak disease; sc_macrophages, CCr2pos;Cx3cr1lo/neg macrophages from spinal cord at peak disease; BMDM_M1/BMDM_M2, bone marrow‐derived macrophages differentiated towards M1 or M2; ECscN and ECscEAE, spinal cord endothelial cells from naïve mice (N) and at peak EAE disease (EAE); SMC, smooth muscle cells from various vessel types (included as positive control to ascertain primer functionality). -

Co-Regulation of Hormone Receptors, Neuropeptides, and Steroidogenic Enzymes 2 Across the Vertebrate Social Behavior Network 3 4 Brent M
bioRxiv preprint doi: https://doi.org/10.1101/435024; this version posted October 4, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 Co-regulation of hormone receptors, neuropeptides, and steroidogenic enzymes 2 across the vertebrate social behavior network 3 4 Brent M. Horton1, T. Brandt Ryder2, Ignacio T. Moore3, Christopher N. 5 Balakrishnan4,* 6 1Millersville University, Department of Biology 7 2Smithsonian Conservation Biology Institute, Migratory Bird Center 8 3Virginia Tech, Department of Biological Sciences 9 4East Carolina University, Department of Biology 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 1 bioRxiv preprint doi: https://doi.org/10.1101/435024; this version posted October 4, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 Running Title: Gene expression in the social behavior network 2 Keywords: dominance, systems biology, songbird, territoriality, genome 3 Corresponding Author: 4 Christopher Balakrishnan 5 East Carolina University 6 Department of Biology 7 Howell Science Complex 8 Greenville, NC, USA 27858 9 [email protected] 10 2 bioRxiv preprint doi: https://doi.org/10.1101/435024; this version posted October 4, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. -

The Role of GABAB(1) Receptor Isoforms in Anxiety and Depression: Genetic and Pharmacological Studies in the Mouse
The Role of GABAB(1) Receptor Isoforms in Anxiety and Depression: Genetic and Pharmacological Studies in the Mouse Inauguraldissertation zur Erlangung der Würde eines Doktors der Philosophie vorgelegt der Philosophisch-Naturwissenschaftlichen Fakultät der Universität Basel von Laura Helen Jacobson aus Neuseeland Basel, 2007 Genehmigt von der Philosophisch-Naturwissenschaftlichen Fakultät auf Antrag von Dissertationsleiterin: Prof. Dr. Anna Wirz-Justice Fakultätsverantwortlicher: Prof. Dr. Heinrich Reichert Korreferentin: Prof. Dr. Carmen Sandi Externe Berater: Dr. John F. Cryan Basel, den 19 Dec 2006 Prof. Dr. Hans-Peter Hauri Dekan For Brian, Liz & Ellis Table of Contents SUMMARY................................................................................................................. 5 ACKNOWLEDGEMENTS .......................................................................................... 8 CHAPTER 1 ............................................................................................................... 9 General Introduction 1.1 The GABAB Receptor.................................................................................................................. 10 1.2 Anxiety and Depression .............................................................................................................. 17 1.3 Mutant Mice and Murine Modelling of Anxiety and Depression.................................................. 18 1.4 GABAB Receptors in Anxiety and Depression ........................................................................... -

Identification of S1PR3 Gene Signature Involved in Survival of Sepsis Patients
Feng et al. BMC Med Genomics (2021) 14:43 https://doi.org/10.1186/s12920-021-00886-2 RESEARCH ARTICLE Open Access Identifcation of S1PR3 gene signature involved in survival of sepsis patients Anlin Feng1, Wenli Ma1, Reem Faraj1, Gabriel T. Kelly1, Stephen M. Black2, Michael B. Fallon1 and Ting Wang1* Abstract Background: Sepsis is a life-threatening complication of infection that rapidly triggers tissue damage in multiple organ systems and leads to multi-organ deterioration. Up to date, prognostic biomarkers still have limitations in pre- dicting the survival of patients with sepsis. We need to discover more prognostic biomarkers to improve the sensitiv- ity and specifcity of the prognosis of sepsis patients. Sphingosine-1-phosphate (S1P) receptor 3 (S1PR3), as one of the S1P receptors, is a prospective prognostic biomarker regulating sepsis-relevant events, including compromised vascular integrity, antigen presentation, and cytokine secretion. Until now, no S1PR3-related prognostic gene signa- tures for sepsis patients have been found. Methods: This study intends to obtain an S1PR3-associated gene signature from whole blood samples to be utilized as a probable prognostic tool for patients with sepsis. Results: We obtained an 18-gene S1PR3-related molecular signature (S3MS) from the intersection of S1PR3-associ- ated genes and survival-associated genes. Numerous important immunity pathways that regulate the progression of sepsis are enriched among our 18 genes. Signifcantly, S3MS functions greatly in both the discovery and validation cohort. Furthermore, we demonstrated that S3MS obtains signifcantly better classifcation performance than random 18-gene signatures. Conclusions: Our results confrm the key role of S1PR3-associated genes in the development of sepsis, which will be a potential prognostic biomarker for patients with sepsis. -

In Vitro Profiling of Orphan G Protein Coupled Receptor (GPCR)
bioRxiv preprint doi: https://doi.org/10.1101/2021.03.10.434788; this version posted March 11, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 1 2 In vitro profiling of orphan G protein coupled receptor (GPCR) 3 constitutive activity 4 5 Lyndsay R. Watkins and Cesare Orlandi * 6 Department of Pharmacology and Physiology, University of Rochester Medical Center, 7 Rochester, NY 14642, USA 8 * Correspondence: [email protected] 9 10 11 12 13 14 15 16 17 18 19 20 Running title: 21 Orphan GPCR constitutive activity 22 23 Keywords: 24 G protein‐coupled receptor (GPCR); constitutive activity; cell signaling; molecular pharmacology. 1 bioRxiv preprint doi: https://doi.org/10.1101/2021.03.10.434788; this version posted March 11, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 25 Abstract 26 Background and Purpose 27 Members of the G protein coupled receptor (GPCR) family are targeted by a significant fraction 28 of the available FDA-approved drugs. However, the physiological role and pharmacological 29 properties of many GPCRs remain unknown, representing untapped potential in drug design. Of 30 particular interest are ~100 less-studied GPCRs known as orphans because their endogenous 31 ligands are unknown. Intriguingly, disease-causing mutations identified in patients, together with 32 animal studies, have demonstrated that many orphan receptors play crucial physiological roles, 33 and thus, represent attractive drug targets. -

Genome-Wide DNA Methylation Analysis Reveals GABBR2 As A
Published OnlineFirst May 10, 2017; DOI: 10.1158/1078-0432.CCR-16-2688 Cancer Therapy: Clinical Clinical Cancer Research Genome-wide DNA Methylation Analysis Reveals GABBR2 as a Novel Epigenetic Target for EGFR 19 Deletion Lung Adenocarcinoma with Induction Erlotinib Treatment Xiaomin Niu1, Fatao Liu2,3, Yi Zhou4, Zhen Zhou1, Daizhan Zhou5, Ting Wang2,3, Ziming Li1, Xiangyun Ye1, Yongfeng Yu1, Xiaoling Weng6,7, Hong Zhang6,7, Junyi Ye6,7, Meilin Liao1, Yun Liu6, Zhiwei Chen1, and Shun Lu1 Abstract Purpose: The past decade has witnessed the rapid development CBFA2T3 and GABBR2, were clearly validated. A same dif- of personalized targeted therapies in lung cancer. It is still unclear ferential methylated region (DMR) between exon 2 and exon whetherepigeneticchangesareinvolvedintheresponsetotyrosine 3ofGABBR2 gene was confirmed consistently in both kinase inhibitor (TKI) treatment in epidermal growth factor recep- patients. GABBR2 was significantly downregulated in EGFR tor (EGFR)-mutated lung cancer. 19 deletion cells, HCC4006 and HCC827, but remained Experimental Design: Methyl-sensitive cut counting sequenc- conserved in EGFR wild-type A549 cells after erlotinib treat- ing (MSCC) was applied to investigate the methylation changes in ment. Upregulation of GABBR2 expression significantly res- paired tissues before and after erlotinib treatment for 42 days with cued erlotinib-induced apoptosis in HCC827 cells. GABBR2 partial response (PR) from stage IIIa (N2) lung adenocarcinoma was significantly downregulated, along with the reduction of patients (N ¼ 2) with EGFR 19 deletion. The Sequenom EpiTYPER S6, p-p70 S6, and p-ERK1/2, demonstrating that GABBR2 assay was used to validate the changed methylated candidate may play an important role in EGFR signaling through the genes. -

Adenylyl Cyclase 2 Selectively Regulates IL-6 Expression in Human Bronchial Smooth Muscle Cells Amy Sue Bogard University of Tennessee Health Science Center
University of Tennessee Health Science Center UTHSC Digital Commons Theses and Dissertations (ETD) College of Graduate Health Sciences 12-2013 Adenylyl Cyclase 2 Selectively Regulates IL-6 Expression in Human Bronchial Smooth Muscle Cells Amy Sue Bogard University of Tennessee Health Science Center Follow this and additional works at: https://dc.uthsc.edu/dissertations Part of the Medical Cell Biology Commons, and the Medical Molecular Biology Commons Recommended Citation Bogard, Amy Sue , "Adenylyl Cyclase 2 Selectively Regulates IL-6 Expression in Human Bronchial Smooth Muscle Cells" (2013). Theses and Dissertations (ETD). Paper 330. http://dx.doi.org/10.21007/etd.cghs.2013.0029. This Dissertation is brought to you for free and open access by the College of Graduate Health Sciences at UTHSC Digital Commons. It has been accepted for inclusion in Theses and Dissertations (ETD) by an authorized administrator of UTHSC Digital Commons. For more information, please contact [email protected]. Adenylyl Cyclase 2 Selectively Regulates IL-6 Expression in Human Bronchial Smooth Muscle Cells Document Type Dissertation Degree Name Doctor of Philosophy (PhD) Program Biomedical Sciences Track Molecular Therapeutics and Cell Signaling Research Advisor Rennolds Ostrom, Ph.D. Committee Elizabeth Fitzpatrick, Ph.D. Edwards Park, Ph.D. Steven Tavalin, Ph.D. Christopher Waters, Ph.D. DOI 10.21007/etd.cghs.2013.0029 Comments Six month embargo expired June 2014 This dissertation is available at UTHSC Digital Commons: https://dc.uthsc.edu/dissertations/330 Adenylyl Cyclase 2 Selectively Regulates IL-6 Expression in Human Bronchial Smooth Muscle Cells A Dissertation Presented for The Graduate Studies Council The University of Tennessee Health Science Center In Partial Fulfillment Of the Requirements for the Degree Doctor of Philosophy From The University of Tennessee By Amy Sue Bogard December 2013 Copyright © 2013 by Amy Sue Bogard.