BULLETINS of AMERICAN PALEONTOLOGY and PALAEONTOGRAPHICA AMERICANA
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

High Influx of Carbon in Walls of Agglutinated Foraminifers During the Permian–Triassic Transition in Global Oceans Galina P
This article was downloaded by: [USGS Libraries] On: 18 February 2015, At: 12:06 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK International Geology Review Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tigr20 High influx of carbon in walls of agglutinated foraminifers during the Permian–Triassic transition in global oceans Galina P. Nestella, Merlynd K. Nestella, Brooks B. Ellwoodb, Bruce R. Wardlawc, Asish R. Basua, Nilotpal Ghoshad, Luu Thi Phuong Lane, Harry D. Rowef, Andrew Hunta, Jonathan H. Tomking & Kenneth T. Ratcliffeh a Department of Earth and Environmental Sciences, University of Texas at Arlington, Arlington, USA b Department of Geology and Geophysics, Louisiana State University, Baton Rouge, USA c Eastern Geology and Palaeoclimate Science Center, United States Geological Survey, Reston, USA Click for updates d Department of Earth and Environmental Sciences, University of Rochester, Rochester, USA e Department of Geomagnetism, Institute of Geophysics, Vietnamese Academy of Science and Technology, Hanoi, Vietnam f Bureau of Economic Geology, University of Texas at Austin, University Station, Austin, USA g School of Earth, Society, and Environment, University of Illinois, Urbana, USA h Chemostrat Inc, Houston, USA Published online: 16 Feb 2015. To cite this article: Galina P. Nestell, Merlynd K. Nestell, Brooks B. Ellwood, Bruce -

Peter H. Griggs
STRATIGRAPHIC SIGNIFICANCE OF FOSSIL POLLEN AND SPORES OF THE CHUCKANUT FORMATION, NORTHWEST WASHINGTON Thosis far the Degree of. M. S. MICHIGAN STATE UNIVERSI'I’Y Peter H. Griggs 1965 ~-.. 'Wfi— wwWWL ‘1. .Illilllflu‘fllllf -Jnl.| IEI‘II‘I'IIIIVFI (II 'llllll‘lll.‘ I, l {III-III! I’lII 'II (III! III, [I , . III! [III [I I'll! (III. (I ABSTRACT STRATIGRAPHIC SIGNIFICANCE OF FOSSIL POLLEN AND SPORES OF THE CHUCKANUT FORMATION, NORTHWEST WASHINGTON by Peter H. Griggs The Chuckanut Formation, a series of strongly folded, terrestrial sandstones and shales, outcrops in northwestern Washington. The rocks have been previously assigned an age of Upper Cretaceous to lower Eocene based upon plant megafossils. The microfossil flora of the standard section along the east shore of Samish Bay was examined in order to determine the relationship of the Chuckanut to the Burrard (middle Eocene) of British Columbia. A zonation and envir- onmental interpretation of the standard section was desired for further refinement of the geological history of Tertiary rocks in the Pacific Northwest. Twenty—two samples, representing 9,H8u feet of section, were examined for palynomorphs. Data were collected for both relative frequency and stratigraphic analysis. Seventy— nine palynomorphs are described. The Samish Bay section is divided into three zones. The zonation is based on changes in the relative frequency f and the stratigraphic range of the palynomorphs. These ’ '_ ‘ V'II -7 r1 ' < P812631 ALL. '.,I I» ) I— (1 0'). OT changes were brought about by changing environmental and climatic conditions during the deposition of the rocks. An age of Paleocene to lower Eocene is assigned to the Samish Bay section. -

CALCARI GRIGI, EARL Y JURASSIC, SOUTHERN ALPS) Si Accettano Manoscritti in Italiano, Inglese E Francese
Direttore Responsabile: Prof. FRANCESCO CoNCONI DIPARTIMENTO DI SCIENZE GEOLOGICHE E PALEONTOLOGICH DELL'UNIVERSITÀ DI FERRARA Aut. Trib. Ferrara n. 36/20.5.53 Biblioteca A.P.A.T. Comitato di redazione della Sezione Scienze della Terra: Proff. A. BosELLINI, C. LORIGA BROGLIO, A. ALBERTI, E. SEMENZA. Gli Annali dell'Università di Ferrara, Sezione Scienze della Terra, vengono inviati in cambio di Riviste scientifiche italiane ed estere; tali riviste sono cedute alle Bibliote che del Dipartimento di Scienze Geologiche e Paleontologiche e dell'Istituto di Mi B 79565 neralogia dell'Università di Ferrara. I lavori da pubblicare (in duplice copia + dischetto) ed ogni comunicazione relativa FAUNAL ASSEMBLAGE AND PALAEOENVIRONMENT alla stampa devono essere inviati in duplice copia a: OF SHALLOW WATER BLACK SHALES Redazione degli Annali, Sezione Se. della T erra, Dipartimento di Scienze Geologiche IN THE TONEZZA AREA e Paleontologiche, C.so Ercole I d'Este 32, 44100 Ferrara. (CALCARI GRIGI, EARL Y JURASSIC, SOUTHERN ALPS) Si accettano manoscritti in italiano, inglese e francese. Gli autori ricevono gratuitamente 50 estratti del loro articolo, senza copertina: BASSI D., BOOMER 1., FUGAGNOLI A., Nome dell'Autore o degli Autori, titolo completo e titolo abbreviato su foglio a parte. LORIGA C., POSENATO R. and WHATLEY R.C. Riassunti: in inglese (ampio se il testo è in italiano) e in italiano (ampio se il testo è in inglese o francese). n riassunto deve evidenziare soprattutto i risultati ottenuti dalla ricerca trala sciando il metodo di lavoro, a meno che questo non rappresenti una novità e parte impo'rtante della pubblicazione. Ringraziamenti, riassunti, key words: con questa successione alla fine del testo e INTRODUCTION prima della bibliografia. -

Jurassic Algae of the Perachora-Peninsula: Biostratigraphical and Paleoecological Implications
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Beiträge zur Paläontologie Jahr/Year: 1994 Band/Volume: 19 Autor(en)/Author(s): Dragastan Ovidiu, Gielisch Hartwig, Richter Detlef K., Grewer Till, Kaziur Thomas, Kube Bärbel, Radusch Christoph Artikel/Article: Jurassic algae of the Perachora-Peninsula: Biostratigraphical and paleoecological implications 49-81 ©Verein zur Förderung der Paläontologie am Institut für Paläontologie, Geozentrum Wien Beitr. Paläont., 19:49-81, Wien 1994 Jurassic algae of the Perachora-Peninsula: Biostratigraphical and paleoecological implications Jurassische Algen der Perachora Halbinsel: Biostratigraphische und paläoökologische Folgerungen by DRAGASTAN, Ovidiu,* GIELISCH, Hartwig**, RICHTER, Detlev K.**, GREWER, TiU**, KAZIUR, Thomas**, KUBE, Bärbel **& RADUSCH, Christoph** DRAGASTAN, O., GIELISCH, H„ RICHTER, D.K., GREWER, T„ KAZIUR, T„ KUBE, B. & RADUSCH, C., 1994. Jurassic algae of the Perachora-Peninsula: Biostratigraphical and paleoecological implications. — Beitr. Palaont., 19:49-81, 9 Figures, 6 Plates, Wien. ceae), Rivularia (Rivulariaceae), Alpinelia graeca n.sp. Contents (Scytonemataceae), Girvanella, Hedstroemia (Porostro- Abstract, Zusammenfassung.................................................. 49 mata) and Microproblematicae. 1. Introduction..............................................................................50 Biostratigraphically, a succession of algae and foramini- 2. Geological setting................................................................. -

VOLCANIC INFLUENCE OVER FLUVIAL SEDIMENTATION in the CRETACEOUS Mcdermott MEMBER, ANIMAS FORMATION, SOUTHWESTERN COLORADO
VOLCANIC INFLUENCE OVER FLUVIAL SEDIMENTATION IN THE CRETACEOUS McDERMOTT MEMBER, ANIMAS FORMATION, SOUTHWESTERN COLORADO Colleen O’Shea A Thesis Submitted to the Graduate College of Bowling Green State University in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE August: 2009 Committee: James Evans, advisor Kurt Panter, co-advisor John Farver ii Abstract James Evans, advisor Volcanic processes during and after an eruption can impact adjacent fluvial systems by high influx rates of volcaniclastic sediment, drainage disruption, formation and failure of natural dams, changes in channel geometry and changes in channel pattern. Depending on the magnitude and frequency of disruptive events, the fluvial system might “recover” over a period of years or might change to some other morphology. The goal of this study is to evaluate the preservation potential of volcanic features in the fluvial environment and assess fluvial system recovery in a probable ancient analog of a fluvial-volcanic system. The McDermott Member is the lower member of the Late Cretaceous - Tertiary Animas Formation in SW Colorado. Field studies were based on a southwest-northeast transect of six measured sections near Durango, Colorado. In the field, 13 lithofacies have been identified including various types of sandstones, conglomerates, and mudrocks interbedded with lahars, mildly reworked tuff, and primary pyroclastic units. Subsequent microfacies analysis suggests the lahar lithofacies can be subdivided into three types based on clast composition and matrix color, this might indicate different volcanic sources or sequential changes in the volcanic center. In addition, microfacies analysis of the primary pyroclastic units suggests both surge and block-and-ash types are present. -

United States
DEPARTMENT OF THE INTERIOR BULLETIN OF THE UNITED STATES ISTo. 146 WASHINGTON GOVERNMENT Pit IN TING OFFICE 189C UNITED STATES GEOLOGICAL SURVEY CHAKLES D. WALCOTT, DI11ECTOK BIBLIOGRAPHY AND INDEX NORTH AMEEICAN GEOLOGY, PALEONTOLOGY, PETEOLOGT, AND MINERALOGY THE YEA.R 1895 FEED BOUGHTON WEEKS WASHINGTON Cr O V E U N M K N T P K 1 N T I N G OFFICE 1890 CONTENTS. Page. Letter of trail smittal...... ....................... .......................... 7 Introduction.............'................................................... 9 List of publications examined............................................... 11 Classified key to tlio index .......................................... ........ 15 Bibliography ............................................................... 21 Index....................................................................... 89 LETTER OF TRANSMITTAL DEPARTMENT OF THE INTEEIOE, UNITED STATES GEOLOGICAL SURVEY, DIVISION OF GEOLOGY, Washington, D. 0., June 23, 1896. SIR: I have the honor to transmit herewith the manuscript of a Bibliography and Index of North American Geology, Paleontology, Petrology, and Mineralogy for the year 1895, and to request that it be published as a bulletin of the Survey. Very respectfully, F. B. WEEKS. Hon. CHARLES D. WALCOTT, Director United States Geological Survey. 1 BIBLIOGRAPHY AND INDEX OF NORTH AMERICAN GEOLOGY, PALEONTOLOGY, PETROLOGY, AND MINER ALOGY FOR THE YEAR 1895. By FRED BOUGHTON WEEKS. INTRODUCTION. The present work comprises a record of publications on North Ameri can geology, paleontology, petrology, and mineralogy for the year 1895. It is planned on the same lines as the previous bulletins (Nos. 130 and 135), excepting that abstracts appearing in regular periodicals have been omitted in this volume. Bibliography. The bibliography consists of full titles of separate papers, classified by authors, an abbreviated reference to the publica tion in which the paper is printed, and a brief summary of the con tents, each paper being numbered for index reference. -

Isopogon & Petrophile
A U S T R AL I A N N A T I V E P L A N T S A S S O C I A T I O N ( A U S T ) Isopogon & Petrophile Study Group Newsletter No. 22 April 2018 ISSN 1445-9493 Website http://anpsa.org.au/iso-petSG/ STUDY GROUP LEADERS/NEWSLETTER EDITORS Catriona Bate & Phil Trickett Email: [email protected] Ph: 0409 789 567 Isopogon teretifolius, Hi Vallee farm, Badgingarra WA, October 2017. See page 10 for our profile of this species. Back issues of the Isopogon and Petrophile Study Group Newsletter are available at http://anpsa.org.au/iso-petSG/IPSG-news.html Isopogon & Petrophile Study Group Newsletter April 2018 1 In this issue Editorial From our members Exchanging cuttings and seed Galls galore Cranbourne I&P Special Collection Painting I. formosus Plant profile – I. teretifolius Plant profile – P. sessilis Dryandra Woodland WA: discovering P. circinata and I. villosus Learnings from Cranbourne Special Collection A seed germination diary: I. anethifolius and P. pulchella Grafting update Seed vs cuttings: P. pedunculata Growing WA natives: experiences in the east and the west Petrophile fossils? Two names discovered In the press Financial report Dear Members, The traditional hot, dry summer reputation of Australia seems to have struck with a vengeance this year. Members have reported little useful rainfall for the entire summer with inland NSW and Victoria/South Australia suffering through one of their driest summers on record. Even here on the normally wet South Coast of NSW, we have had our driest summer since we moved here in 2010. -

Proquest Dissertations
INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from tfie original or copy submitled. Thus, some ttiesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print t>leedthrough, sut)standard margins, and improper alignment can adversely affect reproduction. In the unlikely event tfiat the autfior did not send UMI a complete manuscript and there are missing pages, tfrese will be noted. Also, if unautfiorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced t>y sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overlaps. Photographs included in tfie original manuscript have been reproduced xerographically in this copy. Higher quality 6" x 9" black and white photographic prints are available for any ptwtographs or illustrations appearing in this copy for an additional charge. Contact UMI directly to order. Bell & Howell Information and teaming 300 North zeeb Road, Ann Arbor, Ml 4810B>1346 USA 800-521-0600 UMI* PALEONTOLOGY AND PALEOECOLOGY OF THE NADA MEMBER OF THE BORDEN FORMATION (LOWER MISSISSIPPIAN) IN EASTERN KENTUCKY DISSERTATION Presented in Partial Fullfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University by Aiguo Li, M.S. ***** The Ohio State University 2000 Dissertation Committee: Dr. -

The Yellowstone Paleontological Survey
E PALEONT ON O T LO S G W I O C L A L L E National Y Park The Yellowstone Service Department of the Interior Paleontological Survey SURVEY Vincent L. Santucci Yellowstone Center for Resources National Park Service Yellowstone National Park, Wyoming YCR-NR-98-1 1998 How to cite this document: Santucci, V. L. 1998. The Yellowstone Paleontological Survey. Yellowstone Center for Resources, National Park Service, Yellowstone National Park, Wyoming,YCR-NR-98-1. Current address for Vincent L. Santucci is National Park Service, P.O. Box 592, Kemmerer, WY 83101. The Yellowstone Paleontological Survey To Lt. Col. Luke J. Barnett, III “Uncle by blood, brother in spirit!” Vincent L. Santucci Yellowstone Center for Resources National Park Service Yellowstone National Park, Wyoming YCR-NR-98-1 1998 Table of Contents Introduction .................................................................................................... 1 Stratigraphy .................................................................................................... 4 Fossil Chronology........................................................................................... 6 Taxonomy ..................................................................................................... 12 Localities ...................................................................................................... 15 Interpretation ................................................................................................ 19 Paleontological Resource Management....................................................... -

Biostratigraphy of the Arroyo Penasco Group, Lower Carboniferous (Mississsippian), North-Central New Mexico Augustus K
New Mexico Geological Society Downloaded from: http://nmgs.nmt.edu/publications/guidebooks/25 Biostratigraphy of the Arroyo Penasco Group, Lower Carboniferous (Mississsippian), north-central New Mexico Augustus K. Armstrong and Bernard L. Mamet, 1974, pp. 145-158 in: Ghost Ranch, Siemers, C. T.; Woodward, L. A.; Callender, J. F.; [eds.], New Mexico Geological Society 25th Annual Fall Field Conference Guidebook, 404 p. This is one of many related papers that were included in the 1974 NMGS Fall Field Conference Guidebook. Annual NMGS Fall Field Conference Guidebooks Every fall since 1950, the New Mexico Geological Society (NMGS) has held an annual Fall Field Conference that explores some region of New Mexico (or surrounding states). Always well attended, these conferences provide a guidebook to participants. Besides detailed road logs, the guidebooks contain many well written, edited, and peer-reviewed geoscience papers. These books have set the national standard for geologic guidebooks and are an essential geologic reference for anyone working in or around New Mexico. Free Downloads NMGS has decided to make peer-reviewed papers from our Fall Field Conference guidebooks available for free download. Non-members will have access to guidebook papers two years after publication. Members have access to all papers. This is in keeping with our mission of promoting interest, research, and cooperation regarding geology in New Mexico. However, guidebook sales represent a significant proportion of our operating budget. Therefore, only research papers are available for download. Road logs, mini-papers, maps, stratigraphic charts, and other selected content are available only in the printed guidebooks. Copyright Information Publications of the New Mexico Geological Society, printed and electronic, are protected by the copyright laws of the United States. -
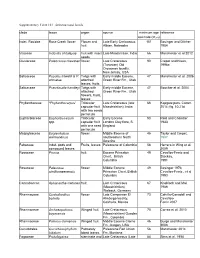
Supplementary Table 10.1. Selected Rosid Fossils Clade Taxon Organ Source Minimum Age Reference Estimate (Mya) Indet
Supplementary Table 10.1. Selected rosid fossils clade taxon organ source minimum age reference estimate (Mya) Indet. Rosidae Rose Creek flower Flower and Late Early Cretaceous 101 Basinger and Dilcher fruit Albian, Nebraska 1984 Vitaceae Indovitis chitaleyae fruit with intact Late Maastrictian, India 66 Manchester et al 2012 seeds Clusiaceae Paleoclusia chevalieri flower Late Cretaceous 90 Crepet and Nixon, (Turonian) Old 1998a Crossman locality, New Jersey, USA Salicaceae Populus tidwellii & P. Twigs with Early middle Eocene, 47 Manchester et al. 2006 wilmatae attached Green River Fm., Utah leaves, fruits Salicaceae Pseudosalix handleyi Twigs with Early middle Eocene, 47 Boucher et al. 2004 attached Green River Fm., Utah flowers, fruits, leaves Phyllanthaceae “Phyllanthocarpus” Trilocular Late Cretaceous (late 66 Kapgate pers. Comm. capsular fruit Maastrichtian), India 2014; fig. 10.21d with two seeds per locule Euphorbiaceae Euphorbiocarpon Trilocular Early Eocene 50 Reid and Chandler spp. capsular fruit London Clay flora, S. 1933 with one seed England per locule Malpighiaceae Eoglandulosa flower Middle Eocene of 45 Taylor and Crepet, warmanensis southeastern North 1987 America Fabaceae Indet. pods and Fruits, leaves Paleocene of Colombia 58 Herrera in Wing et al. compound leaves 2009 Rosaceae Prunus fruit Eocene Princeton 49 Cevallos-Ferriz and Chert, British Stockey, Columbia 1991 Rosaceae Paleorosa flower Middle Eocene 49 Basinger 1976; similkameenensis Princeton Chert, British Cevallos-Ferriz , et al Columbia 1993 Cannabaceae Aphananthe cretacea fruit Late Cretaceous 67 Knobloch and Mai (Maastrichtian) 1986 Walbeck, Germany Rhamnaceae Coahuilanthus flower Late Campanian El 73 Calvillo-Canadell and belinda Almácigo locality, Cevallos- Coahuila, Mexico Ferriz 2007 Rhamnaceae Archaeopaliurus Winged fruit Late Cretaceous 70 Correa et al. -

Ordovician, Silurian, and Middle Devonian Stratigraphy in Northwestern Kentucky and Southern Indiana-Some Reinterpretations
MISCELLANEOUS REPORT NO. 6 ORDOVICIAN, SILURIAN, AND MIDDLE DEVONIAN STRATIGRAPHY IN NORTHWESTERN KENTUCKY AND SOUTHERN INDIANA-SOME REINTERPRETATIONS by James E. Conkin, Barbara M. Conkin, and John Kubacko, Jr. .. • ' .•·"· DIVISION OF GEOLOGICAL SURVEY 4383 FOUNTAIN SQUARE DRIVE COLUMBUS, OHIO 43224-1362 (614) 265-6576 (Voice) (614) 265-6994 {TDD) (614) 447-1918 (FAX) OHIO GEOLOGY ADVISORY COUNCIL Dr. E. Scott Bair, representing Hydrogeology Mr. Mark R. Rowland, representing Environmental Geology Dr. J. Barry Maynard, representing At-Large Citizens Dr. Lon C. Ruedisili, representing Higher Education Mr. Michael T. Puskarich, representing Coal Mr. Gary W. Sitler, representing Oil and Gas Mr. Robert A. Wilkinson, representing Industrial Minerals SCIENTIFIC AND TECHNICAL STAFF OF THE DIVISION OF GEOLOGICAL SURVEY ADMINISTRATION (614) 265-6576 Thomas M. Berg, MS, State Geologist and Division Chief Robert G. Van Hom, MS, Assistant State Geologist and Assistant Division Chief Michael C. Hansen, PhD, Senior Geologist, Ohio Geology Editor, and Geohazards Officer James M . Miller, BA, Fiscal Officer Sharon L. Stone, AD, Executive Secretary REGIONAL GEOLOGY SECTION (614) 265-6597 TECHNICAL PUBLICATIONS SECTION (614) 265-6593 Dennis N. Hull, MS, Geologist Manager and Section Head Merrianne Hackathorn, MS, Geologist and Editor Jean M. Lesher, Typesetting and Printing Technician Paleozoic Geology and Mapping Subsection (614) 265-6473 Edward V. Kuehnle, BA, Cartographer Edward Mac Swinford, MS, Geologist Supervisor Michael R. Lester, BS, Cartographer Glenn E. Larsen, MS, Geologist Robert L. Stewart, Cartographer Gregory A. Schumacher, MS, Geologist Lisa Van Doren, BA, Cartographer Douglas L. Shrake, MS, Geologist Ernie R. Slucher, MS, Geologist PUBLICATIONS CENTER (614) 265-6605 Quaternary Geology and Mapping Subsection (614) 265-6599 Garry E.