Abstract on the Aspen Association of Northern Michigan
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Plant List Bristow Prairie & High Divide Trail
*Non-native Bristow Prairie & High Divide Trail Plant List as of 7/12/2016 compiled by Tanya Harvey T24S.R3E.S33;T25S.R3E.S4 westerncascades.com FERNS & ALLIES Pseudotsuga menziesii Ribes lacustre Athyriaceae Tsuga heterophylla Ribes sanguineum Athyrium filix-femina Tsuga mertensiana Ribes viscosissimum Cystopteridaceae Taxaceae Rhamnaceae Cystopteris fragilis Taxus brevifolia Ceanothus velutinus Dennstaedtiaceae TREES & SHRUBS: DICOTS Rosaceae Pteridium aquilinum Adoxaceae Amelanchier alnifolia Dryopteridaceae Sambucus nigra ssp. caerulea Holodiscus discolor Polystichum imbricans (Sambucus mexicana, S. cerulea) Prunus emarginata (Polystichum munitum var. imbricans) Sambucus racemosa Rosa gymnocarpa Polystichum lonchitis Berberidaceae Rubus lasiococcus Polystichum munitum Berberis aquifolium (Mahonia aquifolium) Rubus leucodermis Equisetaceae Berberis nervosa Rubus nivalis Equisetum arvense (Mahonia nervosa) Rubus parviflorus Ophioglossaceae Betulaceae Botrychium simplex Rubus ursinus Alnus viridis ssp. sinuata Sceptridium multifidum (Alnus sinuata) Sorbus scopulina (Botrychium multifidum) Caprifoliaceae Spiraea douglasii Polypodiaceae Lonicera ciliosa Salicaceae Polypodium hesperium Lonicera conjugialis Populus tremuloides Pteridaceae Symphoricarpos albus Salix geyeriana Aspidotis densa Symphoricarpos mollis Salix scouleriana Cheilanthes gracillima (Symphoricarpos hesperius) Salix sitchensis Cryptogramma acrostichoides Celastraceae Salix sp. (Cryptogramma crispa) Paxistima myrsinites Sapindaceae Selaginellaceae (Pachystima myrsinites) -

Populusspp. Family: Salicaceae Aspen
Populus spp. Family: Salicaceae Aspen Aspen (the genus Populus) is composed of 35 species which contain the cottonwoods and poplars. Species in this group are native to Eurasia/north Africa [25], Central America [2] and North America [8]. All species look alike microscopically. The word populus is the classical Latin name for the poplar tree. Populus grandidentata-American aspen, aspen, bigtooth aspen, Canadian poplar, large poplar, largetooth aspen, large-toothed poplar, poplar, white poplar Populus tremuloides-American aspen, American poplar, aspen, aspen poplar, golden aspen, golden trembling aspen, leaf aspen, mountain aspen, poplar, popple, quaking asp, quaking aspen, quiver-leaf, trembling aspen, trembling poplar, Vancouver aspen, white poplar Distribution Quaking aspen ranges from Alaska through Canada and into the northeastern and western United States. In North America, it occurs as far south as central Mexico at elevations where moisture is adequate and summers are sufficiently cool. The more restricted range of bigtooth aspen includes southern Canada and the northern United States, from the Atlantic coast west to the prairie. The Tree Aspens can reproduce sexually, yielding seeds, or asexually, producing suckers (clones) from their root system. In some cases, a stand could then be composed of only one individual, genetically, and could be many years old and cover 100 acres (40 hectares) or more. Most aspen stands are a mosaic of several clones. Aspen can reach heights of 120 ft (48 m), with a diameter of 4 ft (1.6 m). Aspen trunks can be quite cylindrical, with little taper and few limbs for most of their length. They also can be very crooked or contorted, due to genetic variability. -

Checklist of Common Native Plants the Diversity of Acadia National Park Is Refl Ected in Its Plant Life; More Than 1,100 Plant Species Are Found Here
National Park Service Acadia U.S. Department of the Interior Acadia National Park Checklist of Common Native Plants The diversity of Acadia National Park is refl ected in its plant life; more than 1,100 plant species are found here. This checklist groups the park’s most common plants into the communities where they are typically found. The plant’s growth form is indicated by “t” for trees and “s” for shrubs. To identify unfamiliar plants, consult a fi eld guide or visit the Wild Gardens of Acadia at Sieur de Monts Spring, where more than 400 plants are labeled and displayed in their habitats. All plants within Acadia National Park are protected. Please help protect the park’s fragile beauty by leaving plants in the condition that you fi nd them. Deciduous Woods ash, white t Fraxinus americana maple, mountain t Acer spicatum aspen, big-toothed t Populus grandidentata maple, red t Acer rubrum aspen, trembling t Populus tremuloides maple, striped t Acer pensylvanicum aster, large-leaved Aster macrophyllus maple, sugar t Acer saccharum beech, American t Fagus grandifolia mayfl ower, Canada Maianthemum canadense birch, paper t Betula papyrifera oak, red t Quercus rubra birch, yellow t Betula alleghaniesis pine, white t Pinus strobus blueberry, low sweet s Vaccinium angustifolium pyrola, round-leaved Pyrola americana bunchberry Cornus canadensis sarsaparilla, wild Aralia nudicaulis bush-honeysuckle s Diervilla lonicera saxifrage, early Saxifraga virginiensis cherry, pin t Prunus pensylvanica shadbush or serviceberry s,t Amelanchier spp. cherry, choke t Prunus virginiana Solomon’s seal, false Maianthemum racemosum elder, red-berried or s Sambucus racemosa ssp. -

Recent Declines of Populus Tremuloides in North America Linked to Climate ⇑ James J
Forest Ecology and Management 299 (2013) 35–51 Contents lists available at SciVerse ScienceDirect Forest Ecology and Managemen t journal homepage: www.elsevier.com/locate/foreco Recent declines of Populus tremuloides in North America linked to climate ⇑ James J. Worrall a, , Gerald E. Rehfeldt b, Andreas Hamann c, Edward H. Hogg d, Suzanne B. Marchetti a, Michael Michaelian d, Laura K. Gray c a US Forest Service, Rocky Mountain Region, Gunnison, CO 81230, USA b US Forest Service, Rocky Mountain Research Station, Moscow, ID 83843, USA c University of Alberta, Dept. of Renewable Resources, Edmonton, Alberta, Canada T6G 2H1 d Canadian Forest Service, Northern Forestry Centre, Edmonton, Alberta, Canada T6H 3S5 article info abstract Article history: Populus tremuloides (trembling aspen) recently experie nced extensive crown thinning,branch dieback, Available online 29 January 2013 and mortality across North America. To investigate the role of climate, we developed a range-wide bio- climate model that characterizes clima tic factors controlling distribution ofaspen. We also examined Keywords: indices of moisture stress, insect defoliation and other factors as potential causes of the decline. Historic Decline climate records show that most decline regions experienced exceptionally severe droug htpreceding the Dieback recent episodes. The bioclimate model, driven primarily by maximum summer temperature sand April– Die-off September precipitation, shows that decline tended to occur in marginally suitable habitat, and that cli- Drought matic suitability decreased markedly in the period leading up to decline in almost all decline regions. Climate envelope Climatic niche Other factors, notably multi- year defoliation bytent caterpillars (Malacosoma spp.) and stem damage by fungi and insects, also play a substantial role in decline episodes, and may amplify or prolong the impacts of moisture stress on aspen over large areas. -

Populus Tremula
Populus tremula Populus tremula in Europe: distribution, habitat, usage and threats G. Caudullo, D. de Rigo The Eurasian aspen (Populus tremula L.) is a fast-growing broadleaf tree that is native to the cooler temperate and boreal regions of Europe and Asia. It has an extremely wide range, as a result of which there are numerous forms and subspecies. It can tolerate a wide range of habitat conditions and typically colonises disturbed areas (for example after fire, wind-throw, etc.). It is considered to be a keystone species because of its ecological importance for other species: it has more host-specific species than any other boreal tree. The wood is mainly used for veneer and pulp for paper production as it is light and not particularly strong, although it also has use as a biomass crop because of its fast growth. A number of hybrids have been developed to maximise its vigour and growth rate. Eurasian aspen (Populus tremula L.) is a medium-size, fast-growing tree, exceptionally reaching a height of 30 m1. The Frequency trunk is long and slender, rarely up to 1 m in diameter. The light < 25% branches are rather perpendicular, giving to the crown a conic- 25% - 50% 50% - 75% pyramidal shape. The leaves are 5-7 cm long, simple, round- > 75% ovate, with big wave-shaped teeth2, 3. They flutter in the slightest Chorology Native breeze, constantly moving and rustling, so that trees can often be heard but not seen. In spring the young leaves are coppery-brown and turn to golden yellow in autumn, making it attractive in all vegetative seasons1, 2. -

Poplars and Willows: Trees for Society and the Environment / Edited by J.G
Poplars and Willows Trees for Society and the Environment This volume is respectfully dedicated to the memory of Victor Steenackers. Vic, as he was known to his friends, was born in Weelde, Belgium, in 1928. His life was devoted to his family – his wife, Joanna, his 9 children and his 23 grandchildren. His career was devoted to the study and improve- ment of poplars, particularly through poplar breeding. As Director of the Poplar Research Institute at Geraardsbergen, Belgium, he pursued a lifelong scientific interest in poplars and encouraged others to share his passion. As a member of the Executive Committee of the International Poplar Commission for many years, and as its Chair from 1988 to 2000, he was a much-loved mentor and powerful advocate, spreading scientific knowledge of poplars and willows worldwide throughout the many member countries of the IPC. This book is in many ways part of the legacy of Vic Steenackers, many of its contributing authors having learned from his guidance and dedication. Vic Steenackers passed away at Aalst, Belgium, in August 2010, but his work is carried on by others, including mem- bers of his family. Poplars and Willows Trees for Society and the Environment Edited by J.G. Isebrands Environmental Forestry Consultants LLC, New London, Wisconsin, USA and J. Richardson Poplar Council of Canada, Ottawa, Ontario, Canada Published by The Food and Agriculture Organization of the United Nations and CABI CABI is a trading name of CAB International CABI CABI Nosworthy Way 38 Chauncey Street Wallingford Suite 1002 Oxfordshire OX10 8DE Boston, MA 02111 UK USA Tel: +44 (0)1491 832111 Tel: +1 800 552 3083 (toll free) Fax: +44 (0)1491 833508 Tel: +1 (0)617 395 4051 E-mail: [email protected] E-mail: [email protected] Website: www.cabi.org © FAO, 2014 FAO encourages the use, reproduction and dissemination of material in this information product. -

Poplar Chap 1.Indd
Populus: A Premier Pioneer System for Plant Genomics 1 1 Populus: A Premier Pioneer System for Plant Genomics Stephen P. DiFazio,1,a,* Gancho T. Slavov 1,b and Chandrashekhar P. Joshi 2 ABSTRACT The genus Populus has emerged as one of the premier systems for studying multiple aspects of tree biology, combining diverse ecological characteristics, a suite of hybridization complexes in natural systems, an extensive toolbox of genetic and genomic tools, and biological characteristics that facilitate experimental manipulation. Here we review some of the salient biological characteristics that have made this genus such a popular object of study. We begin with the taxonomic status of Populus, which is now a subject of ongoing debate, though it is becoming increasingly clear that molecular phylogenies are accumulating. We also cover some of the life history traits that characterize the genus, including the pioneer habit, long-distance pollen and seed dispersal, and extensive vegetative propagation. In keeping with the focus of this book, we highlight the genetic diversity of the genus, including patterns of differentiation among populations, inbreeding, nucleotide diversity, and linkage disequilibrium for species from the major commercially- important sections of the genus. We conclude with an overview of the extent and rapid spread of global Populus culture, which is a testimony to the growing economic importance of this fascinating genus. Keywords: Populus, SNP, population structure, linkage disequilibrium, taxonomy, hybridization 1Department of Biology, West Virginia University, Morgantown, West Virginia 26506-6057, USA; ae-mail: [email protected] be-mail: [email protected] 2 School of Forest Resources and Environmental Science, Michigan Technological University, 1400 Townsend Drive, Houghton, MI 49931, USA; e-mail: [email protected] *Corresponding author 2 Genetics, Genomics and Breeding of Poplar 1.1 Introduction The genus Populus is full of contrasts and surprises, which combine to make it one of the most interesting and widely-studied model organisms. -

Auggie Creek Restoration/Fuels Project Threatened, Endangered, and Sensitive Plant Report Darlene Lavelle December 17, 2008
Auggie Creek Restoration/Fuels Project Threatened, Endangered, and Sensitive Plant Report Darlene Lavelle December 17, 2008 Introduction The Seeley Lake Ranger District, Lolo National Forest (LNF), is proposing a restoration project designed to restore forest conditions on approximately 965 acres of Forest Service lands within the Auggie, Seeley, and Mountain Creek drainages. The vegetation treatments are designed to develop a diverse mix of vegetative composition and structure, reduce the risk of bark beetle infestations, and reduce the threat of sustained high intensity wildfire in the wildland-urban interface. Commercial and noncommercial treatments are proposed to reduce stand density, ladder fuels and ground litter, and some dead and down woody debris. Reducing fuels would thereby reduce the risk from insect and disease damage and also the potential for high intensity natural fires around private homes. Fire would also allow an increase of nutrients to plants on the site. Other project proposals include: • Herbicide treatment of weeds along the approximate 12.45 miles of timber haul routes and landings and the approximate 2.37 miles of stored or decommissioned roads mentioned below; • Build about 0.59 miles of temporary road for tree harvest and then decommission these roads. • Store about 1.78 miles of road (close roads to vehicular traffic but keep roads for future use) • Plant western larch and Douglas-fir on about 44 acres within the commercial treatment units to enhance species diversity. • Replace two culverts which are fish barriers, along Swamp Creek and Trail Creek. • Implement additional best management practices (BMPs) involving road drainage at the Morrell Creek Bridge. -

Supplementary Material Table S1. BLAST Database with the Downloaded EST Sequences Involved in the Wood Production Process. in Pa
Supplementary Material Table S1. BLAST database with the downloaded EST sequences involved in the wood production process. In parentheses are the additional keywords used during the search, leading to the resulted sequence collection, presented in the right part of the table No. of the Name of the encoded EST sequences with accession number in NCBI downloaded protein EST sequences GO660523.1, DR749492.1, CO905428.1, CO904299.1, BI975131.1, AW278473.1, BM269700.1, CA936889.1 CA935704.1, BU761220.1, AW428831.1, AW428803.1, AW428739.1, AW428711.1, AJ388815.1, AW163987.1, AI484155.1, AA660372.1, FE969312.1, 14-3-3 like protein FE969243.1, FE969340.1, CN748203.1, CN747244.1, (eudicots) CN746811.1, CN745511.1, CN743519.1, 64 CN747598.1, CN747325.1, CN745584.1, CN745563.1, CN743598.1, CN748506.1, CN748440.1, CN747023.1, GW452329.1, GT662944.1, GW458864.1, GT674095.1, GT674094.1, GT674093.1, GT674092.1, GT712481.1, GT691439.1, GT715099.1, GT704346.1, GT704345.1, GT695499.1, GT695498.1, GT732071.1 GW445847.1, GW450736.1, GW491421.1, GT656115.1, GW431117.1, GW475407.1, GT651591.1, GT651590.1, GT651589.1, GT651588.1, GT660655.1, GW457915.1, GW455608.1, GW457695.1, GW457678.1, GT660154.1, GW430551.1, GW444839.1, GW463204.1, GW462962.1, GW447573.1, GW442110.1, GW441905.1, GW482541.1, GW482491.1, ADP ribosylation factor GT662664.1, GT662618.1, GT668217.1, (eudicots) GW460053.1, GW462758., GW474884.1, 129 GW462640.1, GW462638.1, GW474267.1, GW451892.1, GW441327.1, GW468462.1, GW468212.1, GW451445.1, GW482171.1, GW481960.1, GT664729.1, GW459165.1, GW484383.1, -
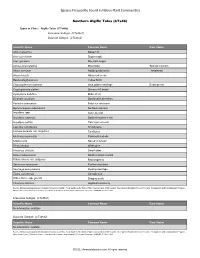
Algific Talus (Cts46)
Species Frequently Found in Native Plant Communities Southern Algific Talus (CTs46) Types in Class: Algific Talus (CTs46a) Limestone Subtype (CTs46a1) Dolomite Subtype (CTs46a2) Scientific Name Column1 Common Name Rare Status Abies balsamea Balsam fir Acer saccharum Sugar maple Acer spicatum Mountain maple Adoxa moschatellina Moschatel Special Concern Allium cernuum Nodding wild onion Threatened Arabis hirsuta Hairy rock cress Betula alleghaniensis Yellow birch Chrysosplenium iowense Iowa golden saxifrage Endangered Cryptogramma stelleri Slender cliff brake Cystopteris bulbifera Bulblet fern Dicentra cucullaria Dutchman's breeches Enemion biternatum False rue anemone Gymnocarpium robertianum Northern oak fern Impatiens spp. touch-me-not Impatiens capensis Spotted touch-me-not Impatiens pallida Pale touch-me-not Laportea canadensis Wood nettle Linnaea borealis var. longiflora Twinflower Mertensia paniculata Panicled bluebells Mitella nuda Naked miterwort Pinus strobus White pine Rhamnus alnifolia Dwarf alder Ribes hudsonianum Northern black currant Rubus idaeus var. strigosus Red raspberry Sambucus racemosa Red-berried elder Saxifraga pensylvanica Swamp saxifrage Taxus canadensis Canada yew Urtica dioica ssp. gracilis Stinging nettle Viburnum trilobum Highbush cranberry Source: Minnesota Department of Natural Resources (2005). Field Guide to the Native Plant Communities of Minnesota: The Eastern Broadleaf Forest Province. Ecological Land Classification Program, Minnesota County Biological Survey, and Natural Heritage and Nongame Research Program. MNDNR St. Paul, MN. Limestone Subtype (CTs46a1) Scientific Name Column1 Common Name Rare Status No information available Dolomite Subtype (CTs46a2) Scientific Name Column1 Common Name Rare Status No information available Source: Minnesota Department of Natural Resources (2005). Field Guide to the Native Plant Communities of Minnesota: The Eastern Broadleaf Forest Province. Ecological Land Classification Program, Minnesota County Biological Survey, and Natural Heritage and Nongame Research Program. -

Kenai National Wildlife Refuge Species List, Version 2018-07-24
Kenai National Wildlife Refuge Species List, version 2018-07-24 Kenai National Wildlife Refuge biology staff July 24, 2018 2 Cover image: map of 16,213 georeferenced occurrence records included in the checklist. Contents Contents 3 Introduction 5 Purpose............................................................ 5 About the list......................................................... 5 Acknowledgments....................................................... 5 Native species 7 Vertebrates .......................................................... 7 Invertebrates ......................................................... 55 Vascular Plants........................................................ 91 Bryophytes ..........................................................164 Other Plants .........................................................171 Chromista...........................................................171 Fungi .............................................................173 Protozoans ..........................................................186 Non-native species 187 Vertebrates ..........................................................187 Invertebrates .........................................................187 Vascular Plants........................................................190 Extirpated species 207 Vertebrates ..........................................................207 Vascular Plants........................................................207 Change log 211 References 213 Index 215 3 Introduction Purpose to avoid implying -

Native Plant List CITY of OREGON CITY 320 Warner Milne Road , P.O
Native Plant List CITY OF OREGON CITY 320 Warner Milne Road , P.O. Box 3040, Oregon City, OR 97045 Phone: (503) 657-0891, Fax: (503) 657-7892 Scientific Name Common Name Habitat Type Wetland Riparian Forest Oak F. Slope Thicket Grass Rocky Wood TREES AND ARBORESCENT SHRUBS Abies grandis Grand Fir X X X X Acer circinatumAS Vine Maple X X X Acer macrophyllum Big-Leaf Maple X X Alnus rubra Red Alder X X X Alnus sinuata Sitka Alder X Arbutus menziesii Madrone X Cornus nuttallii Western Flowering XX Dogwood Cornus sericia ssp. sericea Crataegus douglasii var. Black Hawthorn (wetland XX douglasii form) Crataegus suksdorfii Black Hawthorn (upland XXX XX form) Fraxinus latifolia Oregon Ash X X Holodiscus discolor Oceanspray Malus fuscaAS Western Crabapple X X X Pinus ponderosa Ponderosa Pine X X Populus balsamifera ssp. Black Cottonwood X X Trichocarpa Populus tremuloides Quaking Aspen X X Prunus emarginata Bitter Cherry X X X Prunus virginianaAS Common Chokecherry X X X Pseudotsuga menziesii Douglas Fir X X Pyrus (see Malus) Quercus garryana Garry Oak X X X Quercus garryana Oregon White Oak Rhamnus purshiana Cascara X X X Salix fluviatilisAS Columbia River Willow X X Salix geyeriana Geyer Willow X Salix hookerianaAS Piper's Willow X X Salix lucida ssp. lasiandra Pacific Willow X X Salix rigida var. macrogemma Rigid Willow X X Salix scouleriana Scouler Willow X X X Salix sessilifoliaAS Soft-Leafed Willow X X Salix sitchensisAS Sitka Willow X X Salix spp.* Willows Sambucus spp.* Elderberries Spiraea douglasii Douglas's Spiraea Taxus brevifolia Pacific Yew X X X Thuja plicata Western Red Cedar X X X X Tsuga heterophylla Western Hemlock X X X Scientific Name Common Name Habitat Type Wetland Riparian Forest Oak F.