A New Generic Circumscription in Tribe Lycieae (Solanaceae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Phylogenetic Framework for Evolutionary Study of the Nightshades
Särkinen et al. BMC Evolutionary Biology 2013, 13:214 http://www.biomedcentral.com/1471-2148/13/214 RESEARCH ARTICLE Open Access A phylogenetic framework for evolutionary study of the nightshades (Solanaceae): a dated 1000-tip tree Tiina Särkinen1,2*, Lynn Bohs3, Richard G Olmstead4 and Sandra Knapp1 Abstract Background: The Solanaceae is a plant family of great economic importance. Despite a wealth of phylogenetic work on individual clades and a deep knowledge of particular cultivated species such as tomato and potato, a robust evolutionary framework with a dated molecular phylogeny for the family is still lacking. Here we investigate molecular divergence times for Solanaceae using a densely-sampled species-level phylogeny. We also review the fossil record of the family to derive robust calibration points, and estimate a chronogram using an uncorrelated relaxed molecular clock. Results: Our densely-sampled phylogeny shows strong support for all previously identified clades of Solanaceae and strongly supported relationships between the major clades, particularly within Solanum. The Tomato clade is shown to be sister to section Petota, and the Regmandra clade is the first branching member of the Potato clade. The minimum age estimates for major splits within the family provided here correspond well with results from previous studies, indicating splits between tomato & potato around 8 Million years ago (Ma) with a 95% highest posterior density (HPD) 7–10 Ma, Solanum & Capsicum c. 19 Ma (95% HPD 17–21), and Solanum & Nicotiana c. 24 Ma (95% HPD 23–26). Conclusions: Our large time-calibrated phylogeny provides a significant step towards completing a fully sampled species-level phylogeny for Solanaceae, and provides age estimates for the whole family. -

A Molecular Phylogeny of the Solanaceae
TAXON 57 (4) • November 2008: 1159–1181 Olmstead & al. • Molecular phylogeny of Solanaceae MOLECULAR PHYLOGENETICS A molecular phylogeny of the Solanaceae Richard G. Olmstead1*, Lynn Bohs2, Hala Abdel Migid1,3, Eugenio Santiago-Valentin1,4, Vicente F. Garcia1,5 & Sarah M. Collier1,6 1 Department of Biology, University of Washington, Seattle, Washington 98195, U.S.A. *olmstead@ u.washington.edu (author for correspondence) 2 Department of Biology, University of Utah, Salt Lake City, Utah 84112, U.S.A. 3 Present address: Botany Department, Faculty of Science, Mansoura University, Mansoura, Egypt 4 Present address: Jardin Botanico de Puerto Rico, Universidad de Puerto Rico, Apartado Postal 364984, San Juan 00936, Puerto Rico 5 Present address: Department of Integrative Biology, 3060 Valley Life Sciences Building, University of California, Berkeley, California 94720, U.S.A. 6 Present address: Department of Plant Breeding and Genetics, Cornell University, Ithaca, New York 14853, U.S.A. A phylogeny of Solanaceae is presented based on the chloroplast DNA regions ndhF and trnLF. With 89 genera and 190 species included, this represents a nearly comprehensive genus-level sampling and provides a framework phylogeny for the entire family that helps integrate many previously-published phylogenetic studies within So- lanaceae. The four genera comprising the family Goetzeaceae and the monotypic families Duckeodendraceae, Nolanaceae, and Sclerophylaceae, often recognized in traditional classifications, are shown to be included in Solanaceae. The current results corroborate previous studies that identify a monophyletic subfamily Solanoideae and the more inclusive “x = 12” clade, which includes Nicotiana and the Australian tribe Anthocercideae. These results also provide greater resolution among lineages within Solanoideae, confirming Jaltomata as sister to Solanum and identifying a clade comprised primarily of tribes Capsiceae (Capsicum and Lycianthes) and Physaleae. -

Phylogenetics of the Caprifolieae and Lonicera (Dipsacales)
Systematic Botany (2008), 33(4): pp. 776–783 © Copyright 2008 by the American Society of Plant Taxonomists Phylogenetics of the Caprifolieae and Lonicera (Dipsacales) Based on Nuclear and Chloroplast DNA Sequences Nina Theis,1,3,4 Michael J. Donoghue,2 and Jianhua Li1,4 1Arnold Arboretum of Harvard University, 22 Divinity Ave, Cambridge, Massachusetts 02138 U.S.A. 2Department of Ecology and Evolutionary Biology, Yale University, P.O. Box 208106, New Haven, Conneticut 06520-8106 U.S.A. 3Current Address: Plant Soil, and Insect Sciences, University of Massachusetts at Amherst, Amherst, Massachusetts 01003 U.S.A. 4Authors for correspondence ([email protected]; [email protected]) Communicating Editor: Lena Struwe Abstract—Recent phylogenetic analyses of the Dipsacales strongly support a Caprifolieae clade within Caprifoliaceae including Leycesteria, Triosteum, Symphoricarpos, and Lonicera. Relationships within Caprifolieae, however, remain quite uncertain, and the monophyly of Lonicera, the most species-rich of the traditional genera, and its subdivisions, need to be evaluated. In this study we used sequences of the ITS region of nuclear ribosomal DNA and five chloroplast non-coding regions (rpoB–trnC spacer, atpB–rbcL spacer, trnS–trnG spacer, petN–psbM spacer, and psbM–trnD spacer) to address these problems. Our results indicate that Heptacodium is sister to Caprifolieae, Triosteum is sister to the remaining genera within the tribe, and Leycesteria and Symphoricarpos form a clade that is sister to a monophyletic Lonicera. Within Lonicera, the major split is between subgenus Caprifolium and subgenus Lonicera. Within subgenus Lonicera, sections Coeloxylosteum, Isoxylosteum, and Nintooa are nested within the paraphyletic section Isika. Section Nintooa may also be non-monophyletic. -

Levin and Miller 2005
American Journal of Botany 92(12): 2044±2053. 2005. RELATIONSHIPS WITHIN TRIBE LYCIEAE (SOLANACEAE): PARAPHYLY OF LYCIUM AND MULTIPLE ORIGINS OF GENDER DIMORPHISM1 RACHEL A. LEVIN AND JILL S. MILLER2 Department of Biology, Amherst College, Amherst, Massachusetts 01002 USA We infer phylogenetic relationships among Lycium, Grabowskia, and the monotypic Phrodus microphyllus, using DNA sequence data from the nuclear granule-bound starch synthase gene (GBSSI, waxy) and the chloroplast region trnT-trnF. This is the ®rst comprehensive molecular phylogenetic study of tribe Lycieae (Solanaceae). In addition to providing an understanding of evolutionary relationships, we use the phylogenetic hypotheses to frame our studies of breeding system transitions, ¯oral and fruit evolution, and biogeographical patterns within Lycieae. Whereas Lycium is distributed worldwide, Phrodus and the majority of Grabowskia species are restricted to South America. Tribe Lycieae is strongly supported as monophyletic, but Lycium likely includes both Grabowskia and Phrodus. Results also suggest a single dispersal event from the Americas to the Old World, and frequent dispersal between North and South America. The diversity of fruit types in Lycieae is discussed in light of dispersal patterns and recent work on fruit evolution across Solanaceae. Dimorphic gender expression has been studied previously within Lycium, and results indicate that transitions in sexual expression are convergent, occurring multiple times in North America (a revised estimate from previous studies) and southern Africa. Key words: GBSSI; gender dimorphism; Grabowskia; Lycium; Phrodus; Solanaceae; trnT-trnF; waxy. Tribe Lycieae A.T. Hunziker (Solanaceae) includes Lycium disjunct between the northern and southern hemispheres, since (ca. 80 spp.), Grabowskia (four spp.) and Phrodus (one sp.) Lycium is absent from both the Old and New World tropics. -

Place De La Baie De Goji (Lycium Barbarum L. Solanaceae) Parmi Les Superfruits Actuels : Ses Bienfaits Antioxydants Manon Leroux
Place de la baie de Goji (Lycium barbarum L. Solanaceae) parmi les superfruits actuels : ses bienfaits antioxydants Manon Leroux To cite this version: Manon Leroux. Place de la baie de Goji (Lycium barbarum L. Solanaceae) parmi les superfruits actuels : ses bienfaits antioxydants. Médecine humaine et pathologie. 2014. dumas-01113884 HAL Id: dumas-01113884 https://dumas.ccsd.cnrs.fr/dumas-01113884 Submitted on 6 Feb 2015 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. UNIVERSITE DE ROUEN UFR DE MEDECINE ET DE PHARMACIE Année 2013/2014 N° THESE ƉŽƵƌůĞ/W>KD͛ddKdhZEW,ZD/ Présentée et soutenue publiquement le 3 juillet 2014 par LEROUX Manon Née le 6 avril 1989 à Harfleur Place de la baie de Goji (Lycium barbarum L. Solanaceae) parmi les superfruits actuels : ses bienfaits antioxydants. Président du jury : Mme Elisabeth SEGUIN, Professeur de Pharmacognosie Membres du jury : Mme Marie-Laure GROULT, Maître de Conférences en Botanique Mme Béatrice BLAMPIED ͕WŚĂƌŵĂĐŝĞŶĚ͛ŽĨĨŝĐŝŶĞ >͛hŶŝǀĞƌƐŝƚĠ ĚĞ ZŽƵĞŶ Ğƚ ů͛h&Z ĚĞ DĠĚĞĐŝŶĞ Ğƚ ĚĞ WŚĂƌŵĂĐŝĞ ĚĞ ZŽƵĞŶ Ŷ͛ĞŶƚĞŶĚĞŶƚ ĚŽŶŶĞƌ ĂƵĐƵŶĞ approbation ni improbation aux opinions émises dans cette thèse. Ces opinions sont propres à leurs auteurs. REMERCIEMENTS A Mme Marie->ĂƵƌĞ 'ZKh>d͕ ŵĂ ĚŝƌĞĐƚƌŝĐĞ ĚĞ ƚŚğƐĞ͕ ƉŽƵƌ ŵ͛ĂǀŽŝƌ ĠƉĂƵůĠ e et conseillée tout au long de mon travail. -

Kohn JR (2004) Historical Inferences from the Self-Incompatibility Locus
Chapter 5 What Genealogies of S-alleles Tell Us J.R. Kohn Abstract Drawing on examples from S-RNase-based self-incompatibility (SI), S- locus genealogies are used to infer the demographic history of lineages, the history of mating-system transitions in entire plant families and aspects of the evolution of the S-locus itself. Two lineages of Solanaceae suffered severe restrictions of S-locus diversity evident after millions of years. Broadly shared ancestral S-locus polymorphism is evidence that loss of this form of incompatibility was irreversible in the Solanaceae. Frequent and irreversible loss implies incompatibility is either declining in frequency through time, or that it confers an increased diversification rate relative to self-compatibility (SC). Differences in diversification rate among self-incompatible and self-compatible lineages likely cause the failure of current phylogenetic methods to correctly reconstruct the history of SI. Genealogies also show that origination of new S-RNases rarely occurs within the lifetimes of species. Surprisingly, genealogies of F-box genes purported to provide pollen specificity often do not correspond to those of their cognate S-RNases, indicating we have much to learn about how this system works and evolves. Abbreviations cpDNA DNA from plant plastids GSI Gametophytic SI mya Million years ago SC Self-compatibility SCR/SP11 S-locus cysteine-rich protein (the pollen S-determinant in Brassica) J.R. Kohn Section of Ecology, Behavior and Evolution, Division of Biological Sciences, University of California San Diego, 9500 Gilman Drive, La Jolla CA, 92093–0116 USA, e-mail: [email protected] V.E. Franklin-Tong (ed.) Self-Incompatibility in Flowering Plants – Evolution, Diversity, 103 and Mechanisms. -

Technical Report Series No. 287 Advisory List of Environmental Weeds in Victoria
Advisory list of environmental weeds in Victoria M. White, D. Cheal, G.W. Carr, R. Adair, K. Blood and D. Meagher April 2018 Arthur Rylah Institute for Environmental Research Technical Report Series No. 287 Arthur Rylah Institute for Environmental Research Department of Environment, Land, Water and Planning PO Box 137 Heidelberg, Victoria 3084 Phone (03) 9450 8600 Website: www.ari.vic.gov.au Citation: White, M., Cheal, D., Carr, G. W., Adair, R., Blood, K. and Meagher, D. (2018). Advisory list of environmental weeds in Victoria. Arthur Rylah Institute for Environmental Research Technical Report Series No. 287. Department of Environment, Land, Water and Planning, Heidelberg, Victoria. Front cover photo: Ixia species such as I. maculata (Yellow Ixia) have escaped from gardens and are spreading in natural areas. (Photo: Kate Blood) © The State of Victoria Department of Environment, Land, Water and Planning 2018 This work is licensed under a Creative Commons Attribution 3.0 Australia licence. You are free to re-use the work under that licence, on the condition that you credit the State of Victoria as author. The licence does not apply to any images, photographs or branding, including the Victorian Coat of Arms, the Victorian Government logo, the Department of Environment, Land, Water and Planning logo and the Arthur Rylah Institute logo. To view a copy of this licence, visit http://creativecommons.org/licenses/by/3.0/au/deed.en Printed by Melbourne Polytechnic, Preston Victoria ISSN 1835-3827 (print) ISSN 1835-3835 (pdf)) ISBN 978-1-76077-000-6 (print) ISBN 978-1-76077-001-3 (pdf/online) Disclaimer This publication may be of assistance to you but the State of Victoria and its employees do not guarantee that the publication is without flaw of any kind or is wholly appropriate for your particular purposes and therefore disclaims all liability for any error, loss or other consequence which may arise from you relying on any information in this publication. -

Vegetation Patterns and Dynamics of Renosterveld at Agter-Groeneberg Conservancy, Western Cape, South Africa
Vegetation Patterns and Dynamics of Renosterveld at Agter-Groeneberg Conservancy, Western Cape, South Africa By Benjamin Alan Walton Thesis presented in partial fulfillment of the requirements for the degree of Master of Science at the Stellenbosch University Supervisor Professor Sue J Milton (Department of Conservation Ecology) Co-supervisors A le Roux (CapeNature) Professor L Mucina (Department of Botany and Zoology) April 2006 i Φ Poem “Colour awash over forelands of fertile clay” “When the winters’ cold and grim the Oxalis’s start to brim - they open up. The first feast for bees, in the shrubland short of trees not breeze. Sun’s rays soon last longer in the days: Babianas, Chlorophytums, Geissorhizas, Gladiolius’s, Hesperanthas, Lachenalias, Moraeas and Trachyandras spread their cheerful gaze. Accompanied by annual daisies and bright gladioli filling the air with strong scents of honey - monkey beetles waste no time as they perch upon delicate flowers, lest they are caught in the season’s showers. As if to suggest this is the best nature sends small midge flies to pollinate in jest, and surround mammals to tease their bloody channels. Another month has come and gone - not long now for the raaptol and Micranthus which provide nectar for brown butterflies and painted ladies. Then is the last sequence of bulbs - the fine white-filled fields of chinkerinchees. Grasses’ hour is now soaking up the sun displaying beautifully crafted silhouettes till summers end. As if heaven sent delicate geophytes are still producing their charm, when botanists avoid the midday sun. A brief lapse in displays until the autumn reds begin the seasonal cycles.” Figure a: From left to right: Moraea villosa (Ker Gawl.) Ker Gawl. -

Lycium Barbarum L.) Et Son Utilisation Dans La Santé Humaine
Université de Lille Faculté de pharmacie de Lille Année Universitaire 2018/2019 THESE POUR LE DIPLOME D'ETAT DE DOCTEUR EN PHARMACIE Soutenue publiquement le 10/01/2019 Par M. LEBEAU Pierre-Antoine _____________________________ La baie de Goji (Lycium barbarum L.) et son utilisation dans la santé humaine _____________________________ Membres du jury : Président : DUPONT Fréderic ; Professeur des Universités Assesseur(s) : RIVIERE Cécile ; Maitre de Conférences Membre(s) extérieur(s) : VERMES Philippe ; Docteur en Pharmacie LANGRENE Olivia ; Docteur en Pharmacie 1 Faculté de Pharmacie de Lille 3, rue du Professeur Laguesse - B.P. 83 - 59006 LILLE CEDEX 03.20.96.40.40 - : 03.20.96.43.64 http://pharmacie.univ-lille2.fr Université de Lille Président : Jean-Christophe CAMART Premier Vice-président : Damien CUNY Vice-présidente Formation : Lynne FRANJIÉ Vice-président Recherche : Lionel MONTAGNE Vice-président Relations Internationales : François-Olivier SEYS Directeur Général des Services : Pierre-Marie ROBERT Directrice Générale des Services Adjointe : Marie-Dominique SAVINA Faculté de Pharmacie Doyen : Bertrand DÉCAUDIN Vice-Doyen et Assesseur à la Recherche : Patricia MELNYK Assesseur aux Relations Internationales : : Philippe CHAVATTE Assesseur à la Vie de la Faculté et aux Relations avec le Monde Professionnel : Thomas MORGENROTH Assesseur à la Pédagogie : Benjamin BERTIN Assesseur à la Scolarité : Christophe BOCHU Responsable des Services : Cyrille PORTA Liste des Professeurs des Universités - Praticiens Hospitaliers Civ. NOM Prénom Laboratoire Mme ALLORGE Delphine Toxicologie M. BROUSSEAU Thierry Biochimie M. DÉCAUDIN Bertrand Pharmacie Galénique M. DEPREUX Patrick ICPAL M. DINE Thierry Pharmacie clinique Mme DUPONT-PRADO Annabelle Hématologie M. GRESSIER Bernard Pharmacologie M. LUYCKX Michel Pharmacie clinique M. ODOU Pascal Pharmacie Galénique M. -

Lycium Intricatum Boiss
République Algérienne Démocratique et Populaire Ministère de l’Enseignement Supérieur et de la Recherche Scientifique UNIVERSITE de TLEMCEN Faculté des Sciences de la Nature et de la Vie des Sciences de la Terre et de l’Univers Département d’Ecologie et Environnement MEMOIRE Présenté par Ben allal Hafsa En vue de l’obtention du Diplôme de MASTER En : Ecologie Thème Contribution à l’étude histologique d’une halophyte de l’Oranie : Lycium intricatum Boiss. (Famille : Solanacées) Soutenu le 28 /06 /2018, devant le jury composé de : Président Mme STAMBOULI Hassiba M.C.A Université de Tlemcen Encadreur Mme SARI-ALI Amel M.C.A Université de Tlemcen Examinateur Mme TABTI Nassima M.C.B Université de Tlemcen Remerciements En guise de reconnaissance, je tiens à témoigner mes sincères remerciements à toutes les personnes qui ont contribué de près ou de loin au bon déroulement de mon mémoire de fin d’étude et à l’élaboration de ce modeste travail. Tout d’abord, ce travail ne serait pas aussi riche et n’aurait pas pu voir le jour sans l’aide et l’encadrement de Madame A. Sari-Ali, Maître de conférences A Département d’Ecologie et Environnement nous la remercions pour la qualité de son encadrement exceptionnel, pour sa patience, sa rigueur et sa disponibilité durant notre préparation de ce mémoire. A Madame H. STAMBOULI, Maître de conférences A Département d’Ecologie et Environnement qui nous a fait l’honneur de présider ce jury. A Madame N. TABTI, Maître de conférences B Département d’Ecologie et Environnement , pour avoir accepté d’examiner ce travail. -
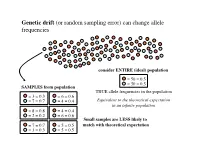
B32-W6-Lecture Copy.Pptx
Genetic drift (or random sampling error) can change allele frequencies! consider ENTIRE (ideal) population! = 50 = 0.5! = 50 = 0.5! SAMPLES from population! TRUE allele frequencies in the population! = 3 = 0.3! = 6 = 0.6! = 7 = 0.7! = 4 = 0.4! Equivalent to the theoretical expectation! in an infinite population! = 8 = 0.8! = 4 = 0.4! = 2 = 0.2! = 6 = 0.6! Small samples are LESS likely to = 7 = 0.7! = 5 = 0.5! match with theoretical expectation! = 3 = 0.3! = 5 = 0.5! Random walks... describe fluctuations in the frequencies of alleles that are neutral with respect to fitness! demes ! Over generations, random sampling error occurs...! During each generation, the large number of zygotes produced is reduced (via mortality that is random with respect to fitness) to some number of breeding individuals! – allele frequencies change (drift) –! What happens to the frequencies of p and q over the long term...?! Alleles drift to fixation in each deme. ! Which allele gets fixed...?! The theory of genetic drift laid out by Sewell Wright during the 1930s and 1940s. Taken up by Motoo Kimura in the 1950s.! Natural selection & genetic drift are the 2 most important causes of evolutionary change in populations. ! The relative importance of each remains controversial.! Genetic drift as a null hypothesis... All alleles @ all loci are subject to genetic drift. The same cannot be said for natural selection.! Evolution by genetic drift...! (1)! Allele frequencies fluctuate at random within a population & eventually one (or another) allele will be fixed.! (2)! The magnitude of change is inversely related to the size of the sample; drift is stronger in smaller samples.! (3)! Drift is NOT repeatable. -

Download Manuscript Manuscript Datphypaper V23 Cleanversion.Docx
Manuscript Clean Version Click here to download Manuscript Manuscript_DatPhyPaper_v23_CleanVersion.docx 1 Phylogenetics of Datureae (Solanaceae), including description of the new genus Trompettia 2 and re-circumscription of the tribe 3 4 Julia Dupin1* and Stacey D. Smith1 5 6 1 Department of Ecology and Evolutionary Biology, University of Colorado Boulder, 1800 7 Colorado Ave, Boulder, CO 80309, USA. 8 * Correspondence author: [email protected] 9 10 Running head: Phylogenetics of Datureae (Solanaceae) 11 12 Abstract 13 14 Datureae G. Don is a tribe in the Solanaceae known for its charismatic large-flowered 15 species (jimsonweeds and angel trumpets). The monophyly of the tribe is well established, but the 16 recent finding that a species previously described in Iochroma (also Solanaceae) belongs in 17 Datureae calls for a reassessment of the tribe’s circumscription. Here we estimated the phylogeny of 18 Datureae, including of all of its 18 species, using three nuclear regions, and incorporated fossil 19 information to estimate divergence times. Based on this phylogeny, we reconstructed the evolution 20 of key aspects of reproductive morphology and life history to identify diagnostic features. Our 21 molecular phylogenetic analyses suggest that the diversification of Datureae began roughly ca. 35 22 Ma, around the beginning of the Andean uplift. Within the tribe, Datura and Brugmansia are 23 monophyletic sister taxa and the misplaced species of Iochroma is sister to the remaining species. 24 Based on our morphological analysis, we describe the latter as a new monotypic genus Trompettia. 25 Ancestral state reconstructions identify diagnostic features for each of the three genera and show a 26 large suite of changes along the Datura branch, including the evolution of erect flowers, capsular 27 fruit and annual life history.