Islands As Model Systems in Ecology and Evolution: Prospects Fifty Years
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Whole-Genome Data Reveal the Complex History of a Diverse Ecological Community
Whole-genome data reveal the complex history of a diverse ecological community Lynsey Bunnefelda,b,1,2, Jack Hearna,1, Graham N. Stonea,3, and Konrad Lohsea,3 aInstitute of Evolutionary Biology, University of Edinburgh, Edinburgh EH9 3FL, Scotland; and bBiological & Environmental Sciences, University of Stirling, Stirling FK9 4LA, Scotland Edited by Craig Moritz, Australian National University, Canberra, ACT, Australia, and accepted by Editorial Board Member Douglas Futuyma May 30, 2018 (received for review January 8, 2018) How widespread ecological communities assemble remains a key Concordant population histories are expected in obligate question in ecology. Trophic interactions between widespread species associations, such as those between plants and special- species may reflect a shared population history or ecological ist pollinators, and these commonly show both coevolution of fitting of local pools of species with very different population his- associated traits and codiversification of the lineages involved tories. Which scenario applies is central to the stability of trophic (6, 7). However, much of terrestrial diversity is found in com- associations and the potential for coevolution between species. munities dominated by less specific interactions between guilds Here we show how alternative community assembly hypotheses of species, for which either codispersal or ecological fitting is can be discriminated using whole-genome data for component a plausible assembly mechanism. These are exemplified by the species and provide a likelihood framework that overcomes cur- rich insect communities associated with temperate trees, in which rent limitations in formal comparison of multispecies histories. widespread herbivores are commonly attacked by a consistent set We illustrate our approach by inferring the assembly history of of parasitoid enemies (8, 9). -

Extinction Patterns, Δ18 O Trends, and Magnetostratigraphy from a Southern High-Latitude Cretaceous–Paleogene Section: Links with Deccan Volcanism
Palaeogeography, Palaeoclimatology, Palaeoecology 350–352 (2012) 180–188 Contents lists available at SciVerse ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Extinction patterns, δ18 O trends, and magnetostratigraphy from a southern high-latitude Cretaceous–Paleogene section: Links with Deccan volcanism Thomas S. Tobin a,⁎, Peter D. Ward a, Eric J. Steig a, Eduardo B. Olivero b, Isaac A. Hilburn c, Ross N. Mitchell d, Matthew R. Diamond c, Timothy D. Raub e, Joseph L. Kirschvink c a University of Washington, Earth and Space Sciences, Box 351310, Seattle WA 98195, United States b CADIC-CONICET, Bernardo Houssay, V9410CAB, Ushuaia, Tierra del Fuego, Argentina c California Institute of Technology, Geological and Planetary Sciences, 1200 E. California Blvd. Pasadena CA 91125, United States d Yale University, Geology & Geophysics, 230 Whitney Ave. New Haven, CT 06511, United States e University of St. Andrews, Department of Earth Sciences, St. Andrews KY16 9AL, UK article info abstract Article history: Although abundant evidence now exists for a massive bolide impact coincident with the Cretaceous–Paleogene Received 19 April 2012 (K–Pg) mass extinction event (~65.5 Ma), the relative importance of this impact as an extinction mechanism is Received in revised form 27 May 2012 still the subject of debate. On Seymour Island, Antarctic Peninsula, the López de Bertodano Formation yields one Accepted 8 June 2012 of the most expanded K–Pg boundary sections known. Using a new chronology from magnetostratigraphy, and Available online 10 July 2012 isotopic data from carbonate-secreting macrofauna, we present a high-resolution, high-latitude paleotemperature record spanning this time interval. -

Estimating Rates of Local Species Extinction, Colonization, and Turnover in Animal Communities
Ecological Applications, 8(4), 1998, pp. 1213±1225 q 1998 by the Ecological Society of America ESTIMATING RATES OF LOCAL SPECIES EXTINCTION, COLONIZATION, AND TURNOVER IN ANIMAL COMMUNITIES JAMES D. NICHOLS,1 THIERRY BOULINIER,2 JAMES E. HINES,1 KENNETH H. POLLOCK,3 AND JOHN R. SAUER1 1U.S. Geological Survey, Biological Resources Division, Patuxent Wildlife Research Center, Laurel, Maryland 20708 USA 2North Carolina Cooperative Fish and Wildlife Research Unit, North Carolina State University, Raleigh, North Carolina 27695 USA 3Institute of Statistics, North Carolina State University, Box 8203, Raleigh, North Carolina 27695-8203 USA Abstract. Species richness has been identi®ed as a useful state variable for conservation and management purposes. Changes in richness over time provide a basis for predicting and evaluating community responses to management, to natural disturbance, and to changes in factors such as community composition (e.g., the removal of a keystone species). Prob- abilistic capture±recapture models have been used recently to estimate species richness from species count and presence±absence data. These models do not require the common assumption that all species are detected in sampling efforts. We extend this approach to the development of estimators useful for studying the vital rates responsible for changes in animal communities over time: rates of local species extinction, turnover, and coloni- zation. Our approach to estimation is based on capture±recapture models for closed animal populations that permit heterogeneity in detection probabilities among the different species in the sampled community. We have developed a computer program, COMDYN, to compute many of these estimators and associated bootstrap variances. -

Ecological Fitting As a Determinant of the Community Structure of Platyhelminth Parasites of Anurans
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Faculty Publications from the Harold W. Manter Laboratory of Parasitology Parasitology, Harold W. Manter Laboratory of 7-2006 Ecological Fitting as a Determinant of the Community Structure of Platyhelminth Parasites of Anurans Daniel R. Brooks University of Toronto, [email protected] Virginia León-Règagnon Nacional Autónoma de México Deborah A. McLennan University of Toronto Derek Zelmer Emporia State University Follow this and additional works at: https://digitalcommons.unl.edu/parasitologyfacpubs Part of the Parasitology Commons Brooks, Daniel R.; León-Règagnon, Virginia; McLennan, Deborah A.; and Zelmer, Derek, "Ecological Fitting as a Determinant of the Community Structure of Platyhelminth Parasites of Anurans" (2006). Faculty Publications from the Harold W. Manter Laboratory of Parasitology. 269. https://digitalcommons.unl.edu/parasitologyfacpubs/269 This Article is brought to you for free and open access by the Parasitology, Harold W. Manter Laboratory of at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Faculty Publications from the Harold W. Manter Laboratory of Parasitology by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Ecology, 87(7) Supplement, 2006, pp. S76-S85 ? 2006 by the Ecological Society of America ECOLOGICAL FITTING AS A DETERMINANT OF THE COMMUNITY STRUCTURE OF PLATYHELMINTH PARASITES OF ANURANS and Daniel R. Brooks,1'5 Virginia Le?n-Regagnon,2 Deborah A. McLennan,3 Derek Zelmer4 1Department of Zoology, University of Toronto, Ontario M5S 3G5 Canada 2Laboratorio de Helmintolog?a, Instituto de Biolog?a, Universidad Nacional Aut?noma de M?xico, C. P. 04510, D. F. M?xico, M?xico of Zoology, University of Toronto, Ontario M5S 3G5 Canada * 3'Department Department of Biological Sciences, Emporia State University, Emporia, Kansas 66801 USA are Abstract. -

Understanding Host-Switching by Ecological Fitting Sabrina B
Virginia Commonwealth University VCU Scholars Compass Biology Publications Dept. of Biology 2015 Understanding Host-Switching by Ecological Fitting Sabrina B. L. Araujo Universidade Federal do Paraná Mariana Pires Braga Universidade Federal do Paraná Daniel R. Brooks Universidade Federal do Paraná See next page for additional authors Follow this and additional works at: http://scholarscompass.vcu.edu/biol_pubs Part of the Biology Commons This is an open access article, free of all copyright, and may be freely reproduced, distributed, transmitted, modified, built upon, or otherwise used by anyone for any lawful purpose. The orkw is made available under the Creative Commons CC0 public domain dedication Downloaded from http://scholarscompass.vcu.edu/biol_pubs/26 This Article is brought to you for free and open access by the Dept. of Biology at VCU Scholars Compass. It has been accepted for inclusion in Biology Publications by an authorized administrator of VCU Scholars Compass. For more information, please contact [email protected]. Authors Sabrina B. L. Araujo, Mariana Pires Braga, Daniel R. Brooks, Salvatore J. Agosta, Eric P. Hoberg, Francisco W. von Hartenthal, and Walter A. Boeger This article is available at VCU Scholars Compass: http://scholarscompass.vcu.edu/biol_pubs/26 RESEARCH ARTICLE Understanding Host-Switching by Ecological Fitting Sabrina B. L. Araujo1,2, Mariana Pires Braga1, Daniel R. Brooks1, Salvatore J. Agosta3, Eric P. Hoberg4, Francisco W. von Hartenthal5, Walter A. Boeger1* 1 Laboratório de Ecologia Molecular e Parasitologia Evolutiva, Universidade Federal do Paraná, Caixa Postal 19073, Curitiba, PR 81531–980, Brazil, 2 Departamento de Física, Universidade Federal do Paraná, Caixa Postal 19044, Curitiba, PR 81531–980, Brazil, 3 Center for Environmental Studies and Department of Biology, Virginia Commonwealth University, Richmond, VA, United States of America, 4 US National Parasite Collection, US Department of Agriculture, Agricultural Research Service, BARC East No. -

Addressing the Mammoth in the Room: the Ethical and Political Implications of De- Extinction
Addressing the Mammoth in the room: The ethical and political implications of de- extinction (Ashlock, 2013) Lowieke Vermeulen (S4374452) Political Science: Political Theory Radboud University, Nijmegen, Netherlands Supervisor: prof. dr. Marcel Wissenburg Date: August 12, 2019 Word count: 23590 1 Table of Contents Chapter 1: Introduction...............................................................................................................3 1.2 Thesis structure............................................................................................................................6 Chapter 2: De-extinction and species selection..........................................................8 2.1 Extinction........................................................................................................................................9 2.2 Approaches to de-extinction.................................................................................................10 2.2.1 Back-breeding.........................................................................................................................10 2.2.2 Cloning.......................................................................................................................................12 2.2.3 Genetic engineering..............................................................................................................12 2.2.4 Mixed approaches..................................................................................................................13 2.3 -

Use of Floral Nectar in Heliconia Stilesii Daniels by Three Species of Hermit Hummingbirds
The Condor89~779-787 0 The Cooper OrnithologicalSociety 1987 ECOLOGICAL FITTING: USE OF FLORAL NECTAR IN HELICONIA STILESII DANIELS BY THREE SPECIES OF HERMIT HUMMINGBIRDS FRANK B. GILL The Academyof Natural Sciences,Philadelphia, PA 19103 Abstract. Three speciesof hermit hummingbirds-a specialist(Eutoxeres aquikz), a gen- eralist (Phaethornissuperciliosus), and a thief (Threnetesruckerz]-visited the nectar-rich flowers of Heliconia stilesii Daniels at a lowland study site on the Osa Peninsula of Costa Rica. Unlike H. pogonanthaCufodontis, a related Caribbean lowland specieswith a less specialized flower, H. stilesii may not realize its full reproductive potential at this site, becauseit cannot retain the services of alternative pollinators such as Phaethornis.The flowers of H. stilesii appear adapted for pollination by Eutoxeres, but this hummingbird rarely visited them at this site. Lek male Phaethornisvisited the flowers frequently in late May and early June, but then abandonedthis nectar sourcein favor of other flowers offering more accessiblenectar. The strong curvature of the perianth prevents accessby Phaethornis to the main nectar chamber; instead they obtain only small amounts of nectar that leaks anteriorly into the belly of the flower. Key words: Hummingbird; pollination; mutualism;foraging; Heliconia stilesii; nectar. INTRODUCTION Ultimately affectedare the hummingbird’s choice Species that expand their distribution following of flowers and patterns of competition among speciationenter novel ecologicalassociations un- hummingbird species for nectar (Stiles 1975, related to previous evolutionary history and face 1978; Wolf et al. 1976; Feinsinger 1978; Gill the challenges of adjustment to new settings, 1978). Use of specificHeliconia flowers as sources called “ecological fitting” (Janzen 1985a). In the of nectar by particular species of hermit hum- case of mutualistic species, such as plants and mingbirds, however, varies seasonally and geo- their pollinators, new ecological settingsmay in- graphically (Stiles 1975). -
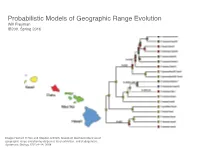
Probabilistic Models of Geographic Range Evolution Will Freyman 10 SYSTEMATIC BIOLOGY VOL
Probabilistic Models of Geographic Range Evolution Will Freyman 10 SYSTEMATIC BIOLOGY VOL. 57 IB200, Spring 2016 Downloaded from http://sysbio.oxfordjournals.org/ at University of California School Law (Boalt Hall) on April 12, 2016 Image: Richard H Ree and Stephen A Smith. Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Systematic Biology, 57(1):4–14, 2008. FIGURE 3. Copyedited by: TRJ MANUSCRIPT CATEGORY: Article 2013 LANDIS ET AL.—BAYESIAN BIOGEOGRAPHY FOR MANY AREAS 3 A) B) There are an infinite number of biogeographic 1234011001 010 011 histories that can explain the observed geographic 011 001 ranges. When calculating the probability of the observed Biogeographic histories on geographic ranges at the tips of the phylogenetic tree, 010 a phylogeny: 5 011 it is unreasonable to condition on a specific history 001 101 of biogeographic change. After all, the past history 101 of biogeographic change is not observable. Instead, 6 111 011 the usual approach is to marginalize over all possible 111 histories of biogeographic change that could give rise 7 to the observed geographic ranges. The standard way 111 Downloaded from 101 to do this is to assume that events of colonization or local extinction occur according to a continuous- 8 101 time Markov chain (Ree et al. 2005). Marginalizing over histories of biogeographic change is accomplished C) D) 011101 010 011 011001 010 011 using two procedures. First, exponentiation of the 010 instantaneous-rate matrix, Q, gives the probability http://sysbio.oxfordjournals.org/ 011 010 011 011 011 011 111 111 001 density of all possible biogeographic changes along a 011 010 branch 111 010 101 011 011 111 011 001 Qt 111 101 011 p(y z t,Q) e− , 111 101 → ; = yz 001 % & 101 111 111 011 where y is the ancestral geographic range, z is the 011 001 current geographic range, and t is the duration of the 001 011 branch on the tree. -

Determining Species Expansion and Extinction Possibilities Using Probabilistic and Graphical Models
Ekológia (Bratislava) Vol. 34, No. 2, p. 111–120, 2015 DOI:10.1515/eko-2015-0012 DetermininG species EXpansion AND EXtinction possibilities UsinG probabilistic AND Graphical models RAJESH CHATURVEDI, P. RAMA CHANDRA PRASAD Lab for Spatial Informatics, International Institute of Information Technology, Gachibowli, Hyderabad - 500032, India; e-mail: [email protected], [email protected] Abstract Chaturvedi R., Prasad R.Ch.: Determining species expansion and extinction possibilities using probabilistic and graphical models. Ekológia (Bratislava), Vol. 34, No. 2, p. 111–120, 2015. Survival of plant species is governed by a number of functions. The participation of each function in species survival and the impact of the contrary behaviour of the species vary from function to function. The probability of extinction of species varies in all such scenarios and has to be calculated separately. Secondly, species follow different patterns of dispersal and localisation at different stages of occupancy state of the site, therefore, the scenarios of competition for resources with climatic shifts leading to deterioration and loss of biodiversity resulting in extinction needs to be studied. Furthermore, most po- ssible deviations of species from climax community states needs to be calculated before species become extinct due to sudden environmental disruption. Globally, various types of anthropogenic disturbances threaten the diversity of biological systems. The impact of these anthropogenic activities needs to be analysed to identify extinction patterns with respect to these activities. All the analyses mentioned abo- ve have been tried to be achieved through probabilistic or graphical models in this study. Key words: biodiversity, disturbances, climate change, endangered, risks, ecological models. -

Mammalian Evolution During the Cretaceous-Tertiary Transition; Evidence Fo~
Acta Palaeontologica Polonica Vol. 28, No. 1-2 pp, 55-61 Warszawa, 1983 Second Symposium on Mesozoic T errestial Ecosystems, Jadwisin 1981 WILLIAM A. CLEMENS MAMMALIAN EVOLUTION DURING THE CRETACEOUS-TERTIARY TRANSITION; EVIDENCE FO~ . GRADUAL, NON-CATASTROPHIC PATTERNS OF BIOTIC CHANGE CLEMENS WILLIAM A. : Mammalian evolution during the Cretaceous-Tertiary transition; ev idence for gradual, non-catastrophic patterns of biotic change. Acta Palaeont. Polonlca, 28, 1-2, 55-61, 1983, Hypotheses Invoking catastrophic causal factors for terminal Cretaceous extinctions call for, 1) Instantaneous, global extinctions, 2) resulting from a single or dominant causal factor. 'c u rr entl y available methods of correlation are not precise enough to test the first element of these hypotheses. Patterns of mammalian extinction and survival In northeastern Montana, USA - as well as records of other organisms - argue against the second element. In Montana most of the lineages of marsupials were decimated. Extinction of multltuber curates was not as severe, while only one lineage of eutherlans appears to have become extinct. Groups that apparently Immigrated Into northeastern Montana just prior to the end of the Cretaceous had a greater probability of survival than the residents. The prObability of extinction was Independent of Individual body size. Different patterns of mammalian extinction In different environments also appear to defy explanation In terms of one common, catastrophic causal factor. Key w 0 r d s: Mammalia, Cretaceous, Tertiary, extinction, Montana, USA . Wltltam A . Clemens. Department Of Paleontology. University 01 Callfornla. Berkeley, Callfornla. 94720. USA. Recei ved: September 1981. INTRODUCTION Scientific and popular publications continue to be flooded with articles concerning the causes of the extinction of dinosaurs and other organisms used to mark the end of the Cretaceous. -

Evidence for Local and Global Redox Conditions at an Early Ordovician (Tremadocian) Mass Extinction ∗ ∗∗ Cole T
Earth and Planetary Science Letters 481 (2018) 125–135 Contents lists available at ScienceDirect Earth and Planetary Science Letters www.elsevier.com/locate/epsl Evidence for local and global redox conditions at an Early Ordovician (Tremadocian) mass extinction ∗ ∗∗ Cole T. Edwards a,b, , David A. Fike b, Matthew R. Saltzman c, Wanyi Lu d, Zunli Lu d, a Department of Geological and Environmental Sciences, Appalachian State University, Boone, NC 28608, USA b Department of Earth and Planetary Sciences, Washington University, St. Louis, MO 63130, USA c School of Earth Sciences, The Ohio State University, Columbus, OH 43214, USA d Department of Earth Sciences, Syracuse University, Syracuse, NY 13244, USA a r t i c l e i n f o a b s t r a c t Article history: Profound changes in environmental conditions, particularly atmospheric oxygen levels, are thought to be Received 29 May 2017 important drivers of several major biotic events (e.g. mass extinctions and diversifications). The early Received in revised form 20 September Paleozoic represents a key interval in the oxygenation of the ocean–atmosphere system and evolution 2017 of the biosphere. Global proxies (e.g. carbon (δ13C) and sulfur (δ34S) isotopes) are used to diagnose Accepted 1 October 2017 potential changes in oxygenation and infer causes of environmental change and biotic turnover. The Available online xxxx Cambrian–Ordovician contains several trilobite extinctions (some are apparently local, but others are Editor: D. Vance 13 34 globally correlative) that are attributed to anoxia based on coeval positive δ C and δ S excursions. These Keywords: extinction and excursion events have yet to be coupled with more recently developed proxies thought Ordovician to be more reflective of local redox conditions in the water column (e.g. -

The Indirect Paths to Cascading Effects of Extinctions in Mutualistic Networks
The indirect paths to cascading effects of extinctions in mutualistic networks Item Type Article Authors Pires, Mathias M.; O'Donnell, James L.; Burkle, Laura A.; Díaz- Castelazo, Cecilia; Hembry, David H.; Yeakel, Justin D.; Newman, Erica A.; Medeiros, Lucas P.; de Aguiar, Marcus A. M.; Guimarães, Paulo R. Jr Citation Pires, M. M., O'Donnell, J. L., Burkle, L. A., DíazCastelazo, C., Hembry, D. H., Yeakel, J. D., ... & Guimarães Jr, P. R. (2020). The indirect paths to cascading effects of extinctions in mutualistic networks. Ecology, e03080. DOI 10.1002/ecy.3080 Publisher WILEY Journal ECOLOGY Rights © 2020 by the Ecological Society of America. Download date 05/10/2021 23:12:03 Item License http://rightsstatements.org/vocab/InC/1.0/ Version Final published version Link to Item http://hdl.handle.net/10150/648146 Reports Ecology, 101(7), 2020, e03080 © 2020 by the Ecological Society of America The indirect paths to cascading effects of extinctions in mutualistic networks 1,11 2 3 4 MATHIAS M. PIRES , JAMES L. O'DONNELL, LAURA A. BURKLE , CECILIA DIAZ-CASTELAZO , 5,6 7 6 8 DAVID H. HEMBRY , JUSTIN D. YEAKEL , ERICA A. NEWMAN , LUCAS P. M EDEIROS , 9 10 MARCUS A. M. DE AGUIAR , AND PAULO R. GUIMARAES~ JR. 1Departamento de Biologia Animal, Instituto de Biologia, Universidade Estadual de Campinas, Campinas,13.083-862, Sao~ Paulo, Brazil 2School of Marine and Environmental Affairs, University of Washington, Seattle, WA 98105, Washington, USA 3Department of Ecology, Montana State University, Bozeman, MT 59717, Montana, USA 4Red de Interacciones