Bahinia Pondaungensis, an Alleged Early Anthropoid
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The World at the Time of Messel: Conference Volume
T. Lehmann & S.F.K. Schaal (eds) The World at the Time of Messel - Conference Volume Time at the The World The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment and the History of Early Primates 22nd International Senckenberg Conference 2011 Frankfurt am Main, 15th - 19th November 2011 ISBN 978-3-929907-86-5 Conference Volume SENCKENBERG Gesellschaft für Naturforschung THOMAS LEHMANN & STEPHAN F.K. SCHAAL (eds) The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates 22nd International Senckenberg Conference Frankfurt am Main, 15th – 19th November 2011 Conference Volume Senckenberg Gesellschaft für Naturforschung IMPRINT The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates 22nd International Senckenberg Conference 15th – 19th November 2011, Frankfurt am Main, Germany Conference Volume Publisher PROF. DR. DR. H.C. VOLKER MOSBRUGGER Senckenberg Gesellschaft für Naturforschung Senckenberganlage 25, 60325 Frankfurt am Main, Germany Editors DR. THOMAS LEHMANN & DR. STEPHAN F.K. SCHAAL Senckenberg Research Institute and Natural History Museum Frankfurt Senckenberganlage 25, 60325 Frankfurt am Main, Germany [email protected]; [email protected] Language editors JOSEPH E.B. HOGAN & DR. KRISTER T. SMITH Layout JULIANE EBERHARDT & ANIKA VOGEL Cover Illustration EVELINE JUNQUEIRA Print Rhein-Main-Geschäftsdrucke, Hofheim-Wallau, Germany Citation LEHMANN, T. & SCHAAL, S.F.K. (eds) (2011). The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates. 22nd International Senckenberg Conference. 15th – 19th November 2011, Frankfurt am Main. Conference Volume. Senckenberg Gesellschaft für Naturforschung, Frankfurt am Main. pp. 203. -

From the Gulf Coastal Plain
Bull. Fla. Mus. Nat. Hist. (2005) 45(4): 355-361 355 EKGMOWECHASHALA (MAMMALIA, ?PRIMATES) FROM THE GULF COASTAL PLAIN L. Barry Albright III1 A single, small, water-worn tooth from the “middle” Arikareean Toledo Bend Local Fauna of the Gulf Coastal Plain closely resembles the lower fourth premolar of the questionable primate Ekgmowechashala. The only known species of the genus, Ekgmowechashala philotau Macdonald (1963), was originally recovered from the early Arikareean Sharps Formation of South Dakota, but is also known from similar aged strata of the John Day Formation, Oregon. Although the Toledo Bend specimen differs somewhat in morphology from the p4 of E. philotau, a new species is not named in this report because of such limited material and because the specimen is incomplete. Unfortunately, the specimen does not provide information that helps clarify current arguments regarding the affinities of Ekgmowechashala with primates or plagiomenids. It does, however, provide (1) a temporal range extension for the genus to the early late Arikareean, or about four million years younger than previously known, and (2) a geographic extension east and considerably south of its prior distribution. If Ekgmowechashala is ultimately deter- mined to belong to the Primates, then the Toledo Bend species would become the last known North American representative of the order. Key Words: Ekgmowechashala; Primates; Plagiomenidae; Texas Gulf Coastal Plain; Arikareean INTRODUCTION brate paleontology collections of the Louisiana State Uni- In 1990, while screen-washing matrix from the “middle” versity Museum of Geoscience (LSUMG V). Arikareean Toledo Bend site in the Fleming Formation of easternmost Texas (Albright 1994, 1996, 1998a, 1998b, BACKGROUND 1999), a small, unusual, water-worn tooth was recov- Ekgmowechashala philotau is a small “enigmatic late ered that differed considerably from the site’s more com- Oligocene mammal” (McKenna, 1990:226) heretofore mon and readily identifiable rodent teeth. -

Convergent Evolution of Olfactory and Thermoregulatory Capacities in Small Amphibious Mammals
Convergent evolution of olfactory and thermoregulatory capacities in small amphibious mammals Quentin Martineza,1, Julien Clavelb,c, Jacob A. Esselstynd,e, Anang S. Achmadif, Camille Grohég,h, Nelly Piroti,j, and Pierre-Henri Fabrea,k aInstitut des Sciences de l’Évolution de Montpellier (ISEM), CNRS, Institut de recherche pour le développement (IRD), Université de Montpellier (UM), UMR 5554, 34095 Montpellier, France; bDepartment of Life Sciences, The Natural History Museum, SW7 5DB London, United Kingdom; cUniv. Lyon Laboratoire d’Ecologie des Hydrosystèmes Naturels et Anthropisés, UMR CNRS 5023, Université Claude Bernard Lyon 1, École Nationale des Travaux Publics de l’État (ENTPE), F‐69622 Villeurbanne, Cedex, France; dMuseum of Natural Science, Louisiana State University, Baton Rouge, LA 70803; eDepartment of Biological Sciences, Louisiana State University, Baton Rouge, LA 70803; fMuseum Zoologicum Bogoriense, Research Center for Biology, Indonesian Institute of Sciences (LIPI), 16911 Cibinong, Indonesia; gDivision of Paleontology, American Museum of Natural History, New York, NY 10024; hLaboratoire Paléontologie Évolution Paléoécosystèmes Paléoprimatologie (PALEVOPRIM, UMR 7262, CNRS-Institut écologie et environnement [INEE]), Université de Poitiers, 86073 Poitiers, Cedex 9, France; iInstitut de Recherche en Cancérologie de Montpellier (IRCM), INSERM, U1194 UM, Institut du Cancer de Montpellier (ICM), F-34298 Montpellier, Cedex 5, France; jRéseau d’Histologie Expérimentale de Montpellier, UMS3426 CNRS-US009 INSERM-UM, 34298 Montpellier, France; and kMammal Section, Department of Life Sciences, The Natural History Museum, SW7 5DB London, United Kingdom Edited by David B. Wake, University of California, Berkeley, CA, and approved February 28, 2020 (received for review October 11, 2019) Olfaction and thermoregulation are key functions for mammals. The partitioning has been documented in histological, airflow dynamic, former is critical to feeding, mating, and predator avoidance behaviors, and performance test studies (9–13). -

8. Primate Evolution
8. Primate Evolution Jonathan M. G. Perry, Ph.D., The Johns Hopkins University School of Medicine Stephanie L. Canington, B.A., The Johns Hopkins University School of Medicine Learning Objectives • Understand the major trends in primate evolution from the origin of primates to the origin of our own species • Learn about primate adaptations and how they characterize major primate groups • Discuss the kinds of evidence that anthropologists use to find out how extinct primates are related to each other and to living primates • Recognize how the changing geography and climate of Earth have influenced where and when primates have thrived or gone extinct The first fifty million years of primate evolution was a series of adaptive radiations leading to the diversification of the earliest lemurs, monkeys, and apes. The primate story begins in the canopy and understory of conifer-dominated forests, with our small, furtive ancestors subsisting at night, beneath the notice of day-active dinosaurs. From the archaic plesiadapiforms (archaic primates) to the earliest groups of true primates (euprimates), the origin of our own order is characterized by the struggle for new food sources and microhabitats in the arboreal setting. Climate change forced major extinctions as the northern continents became increasingly dry, cold, and seasonal and as tropical rainforests gave way to deciduous forests, woodlands, and eventually grasslands. Lemurs, lorises, and tarsiers—once diverse groups containing many species—became rare, except for lemurs in Madagascar where there were no anthropoid competitors and perhaps few predators. Meanwhile, anthropoids (monkeys and apes) emerged in the Old World, then dispersed across parts of the northern hemisphere, Africa, and ultimately South America. -

Early Eocene Primates from Gujarat, India
ARTICLE IN PRESS Journal of Human Evolution xxx (2009) 1–39 Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Early Eocene Primates from Gujarat, India Kenneth D. Rose a,*, Rajendra S. Rana b, Ashok Sahni c, Kishor Kumar d, Pieter Missiaen e, Lachham Singh b, Thierry Smith f a Johns Hopkins University School of Medicine, Baltimore, Maryland 21205, USA b H.N.B. Garhwal University, Srinagar 246175, Uttarakhand, India c Panjab University, Chandigarh 160014, India d Wadia Institute of Himalayan Geology, Dehradun 248001, Uttarakhand, India e University of Ghent, B-9000 Ghent, Belgium f Royal Belgian Institute of Natural Sciences, B-1000 Brussels, Belgium article info abstract Article history: The oldest euprimates known from India come from the Early Eocene Cambay Formation at Vastan Mine Received 24 June 2008 in Gujarat. An Ypresian (early Cuisian) age of w53 Ma (based on foraminifera) indicates that these Accepted 8 January 2009 primates were roughly contemporary with, or perhaps predated, the India-Asia collision. Here we present new euprimate fossils from Vastan Mine, including teeth, jaws, and referred postcrania of the Keywords: adapoids Marcgodinotius indicus and Asiadapis cambayensis. They are placed in the new subfamily Eocene Asiadapinae (family Notharctidae), which is most similar to primitive European Cercamoniinae such as India Donrussellia and Protoadapis. Asiadapines were small primates in the size range of extant smaller Notharctidae Adapoidea bushbabies. Despite their generally very plesiomorphic morphology, asiadapines also share a few derived Omomyidae dental traits with sivaladapids, suggesting a possible relationship to these endemic Asian adapoids. In Eosimiidae addition to the adapoids, a new species of the omomyid Vastanomys is described. -

Paleo-The Story of Life
PALEO: THE STORY OF LIFE Life on Earth has not always existed as it currently does. The fact that life began on Earth in the first place is miraculous due to the environmental factors needed for its beginnings and sustainability. The relentless pursuit of life over billions of years from small living molecules to complex creatures roaming, flying and swimming throughout the Earth has culminated into the current state of life’s existence as we know it on the planet we call home. Paleo: The Story of Life is a 3,000-square-foot exhibit, spanning 4.6 billion years in scope. The exhibit presents casts of 128 rare fossils, including Lucy, Archaeopteryx and T rex, among many others. Drawn from the world’s foremost fossil collections, the Paleo exhibit showcases casts of rare fossils from the Americas, Europe, Asia, Africa and Australia – skeletons, skulls, claws and eggs gathered from prestigious museums, including the Smithsonian Institution, American Museum of Natural History, Royal Ontario Museum and Carnegie Museum, among others. Rarely available for viewing outside of their respective museums, these compelling artifacts are presented exclusively in Paleo: The Story of Life. Fossils range from the earliest invertebrate marine life through the Triassic, Jurassic and Cretaceous dinosaurs to mammals and prehistoric humans. Paleo: The Story of Life explores the comprehensive story of prehistoric life on Earth. The Paleo exhibit is a visiting exhibit and will be on display through Thursday, May 31, 2018. It is located in the Horowitz Traveling Exhibit Area. The MOST presents Paleo: The Story of Life in association with the International Museum Institute, Inc. -

Cranial Remains of an Eocene Tarsier
Cranial remains of an Eocene tarsier James B. Rossie*†, Xijun Ni‡, and K. Christopher Beard§ *Department of Anthropology, Stony Brook University, Stony Brook, NY 11794; ‡Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, P.O. Box 643, Beijing 100044, China; §Section of Vertebrate Paleontology, Carnegie Museum of Natural History, 4400 Forbes Avenue, Pittsburgh, PA 15213 Edited by Alan Walker, Pennsylvania State University, University Park, PA, and approved January 30, 2006 (received for review October 28, 2005) The phylogenetic position of tarsiers relative to anthropoids and Paleogene omomyids remains a subject of lively debate that lies at the center of research into anthropoid origins. Omomyids have long been regarded as the nearest relatives of tarsiers, but a sister group relationship between anthropoids and tarsiers has also been proposed. These conflicting phylogenetic reconstructions rely heavily on comparisons of cranial anatomy, but until now, the fossil record of tarsiers has been limited to a single jaw and several isolated teeth. In this article, we describe cranial material of a fossil tarsiid from the middle-Eocene Shanghuang fissure-fillings in southern Jiangsu Province, China. This facial fragment, which is allocated to Tarsius eocaenus, is virtually identical to the corre- sponding anatomy in living tarsiers and differs substantially from that of early anthropoids such as Bahinia, Phenacopithecus, and Parapithecus. This new specimen indicates that tarsiers already possessed greatly enlarged orbits and a haplorhine oronasal con- figuration by the time they are first documented in the fossil record during the middle Eocene. China ͉ primate ͉ tarsiid ack of consensus regarding the interrelationships among Lanthropoids, tarsiids, and omomyids is largely due to con- tradictory similarities among the three groups in cranial anat- omy. -

Evidence for an Asian Origin of Stem Anthropoids
Evidence for an Asian origin of stem anthropoids Richard F. Kay1 Department of Evolutionary Anthropology, Duke University, Durham, NC 27708-03083 n PNAS, Chaimanee et al. (1) report Genetic, embryological, and anatomical We are not there yet. Recent comprehen- a previously undescribed species of evidence demonstrates that the sister sive phylogenetic analyses stemming from I primate, Afrasia, from the late Middle group of Anthropoidea is the south Asian virtually the same datasets yield somewhat Eocene of Burma. They identify tarsier, with the two forming the crown different cladograms that reflect sensitivity Afrasia as the sister taxon to the African group Haplorhini (6–8). The other clade of to which taxa are included in the analysis genus Afrotarsius but slightly more primitive extant primates is the lemurs and lorises, and which sets of analytical assumptions are than it and allied with stem Anthropoidea called Strepsirrhini. Eocene Holarctic selected. Pertinent to the biogeographic of south Asia. Anthropoidea is the taxo- Omomyoidea are generally considered as conclusions of Chaimanee et al. (1), Seiffert nomic group that today includes New and stem haplorhines, although the precise et al. (11) conclude that Afrotarsius cha- Old World monkeys, apes, and humans. If relationship of omomyoids to tarsiers and trathi [a younger species than the one that upheld, the biogeographic significance of anthropoids is uncertain (8). Tarsius often Chaimanee et al. (1) describe] is an African these results is profound: If Afrasia and is considered to be a relictual omomyoid tarsioid. If Seiffert et al. (11) are correct, Afrotarsius are as closely related as Chai- in south Asia. -

A Fossil Primate of Uncertain Affinities from the Earliest Late Eocene of Egypt
A fossil primate of uncertain affinities from the earliest late Eocene of Egypt Erik R. Seifferta,1, Elwyn L. Simonsb,1, Doug M. Boyerc, Jonathan M. G. Perryd, Timothy M. Ryane, and Hesham M. Sallamf aDepartment of Anatomical Sciences, Stony Brook University, Stony Brook, NY 11794-8081; bDivision of Fossil Primates, Duke Lemur Center, Durham, NC 27705; cDepartment of Ecology and Evolution, Stony Brook University, Stony Brook, NY 11794-5245; dDepartment of Anatomy, Midwestern University, Downers Grove, IL 60515; eDepartment of Anthropology, Pennsylvania State University, University Park, PA 16802; and fDepartment of Earth Sciences, University of Oxford, Oxford OX1 3PR, United Kingdom Contributed by Elwyn L. Simons, February 18, 2010 (sent for review January 5, 2010) Paleontological work carried out over the last 3 decades has es- as caenopithecines are likely derived from an independent colo- tablished that three major primate groups were present in the Eocene nization from Europe or Asia. The timing of anthropoids’ arrival in of Africa—anthropoids, adapiforms, and advanced strepsirrhines. Afro-Arabia is unclear, but the presence of multiple anthropoid Here we describe isolated teeth of a previously undocumented pri- species at BQ-2 supports a colonization in the late middle Eocene mate from the earliest late Eocene (≈37 Ma) of northern Egypt, Nos- or earlier. mips aenigmaticus, whose phylogenetic placement within Primates is Here we describe a rare and enigmatic primate species from BQ-2 unclear. Nosmips is smaller than the sympatric adapiform Afradapis that, unlike other primates from theEoceneofAfrica,doesnotfit but is considerably larger than other primate taxa known from the unambiguously into either Anthropoidea or Strepsirrhini on the basis same paleocommunity. -
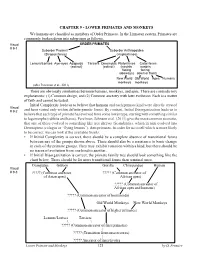
CHAPTER 9 - LOWER PRIMATES and MONKEYS We Humans Are Classified As Members of Order Primates
CHAPTER 9 - LOWER PRIMATES AND MONKEYS We humans are classified as members of Order Primates. In the Linnaean system, Primates are commonly broken down into subgroups as follows. Visual ORDER PRIMATES # 9-1 Suborder Prosimii Suborder Anthropoidea (Strepsirrhines) (Haplorrhines) Lemurs/Lorises Aye-ayes Adapoids Tarsiers Omomyids Platyrrhines Catarrhines (extinct) (extinct) (nostrils nostrils facing facing sideways) down or front) New World Old World Apes Humans monkeys monkeys (after Perelman et al., 2011) There are obviously similarities between humans, monkeys, and apes. There are contradictory explanations: (1) Common design, and (2) Common ancestry with later evolution. Each is a matter of faith and cannot be tested. Initial Complexity leads us to believe that humans and each primate kind were directly created Visual # 9-2 and have varied only within definite genetic limits. By contrast, Initial Disorganization leads us to believe that each type of primate has evolved from some lower type, starting with something similar to lagomorphs (rabbits and hares). Perelman, Johnson et al. (2011) give the most common scenario, that one of these evolved to something like tree shrews (Scandentia), which in turn evolved into Dermoptera (colugos or “flying lemurs”), then primates. In order for us to tell which is more likely to be correct, we can look at the available fossils. • If Initial Complexity is correct, there should be a complete absence of transitional forms between any of the groups shown above. There should also be a resistance to basic change in each of the primate groups. They may exhibit variation within a kind, but there should be no traces of evolution from one kind to another. -

The First Virtual Endocasts of North American Adapiform Primates
THE FIRST VIRTUAL ENDOCASTS OF NORTH AMERICAN ADAPIFORM PRIMATES By ARIANNA R. HARRINGTON A THESIS PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF THE MASTER OF SCIENCE UNIVERSITY OF FLORIDA 2015 © 2015 Arianna R. Harrington ACKNOWLEDGEMENTS I would like to full-heartedly thank Dr. Jonathan Bloch of the Florida Museum of Natural History for serving as my primary advisor and committee chair, and Dr. Bruce MacFadden of the Florida Museum of Natural History and Dr. David Daegling of the Department of Anthropology at the University of Florida for serving on my committee. I would like to thank all three of my committee members for their time and helpful suggestions with my project and this manuscript. I would also like to thank Dr. Mary Silcox of the University of Toronto, Scarborough, for her help, advice, and training with regards to the reconstruction of virtual endocasts, Dr. Douglas Boyer of Duke University for help acquiring, scanning, and facilitating access to specimens used for this study, and Gabriel Yapuncich of Duke University for helpful conversations about body mass estimation. I would also like to thank my family and friends for their support during the course of my education. 3 TABLE OF CONTENTS page ACKNOWLEDGEMENTS .............................................................................................................3 LIST OF TABLES ...........................................................................................................................6 -

The Tempo of Trophic Evolution in Small-Bodied Primates 2 3 4 Running Title: Trophic Evolution in Small-Bodied Primates 5 6 7 Jeremiah E
bioRxiv preprint doi: https://doi.org/10.1101/2020.03.17.996207; this version posted March 20, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-ND 4.0 International license. 1 1 The tempo of trophic evolution in small-bodied primates 2 3 4 Running title: Trophic evolution in small-bodied primates 5 6 7 Jeremiah E. Scott 8 Department of Medical Anatomical Sciences 9 College of Osteopathic Medicine of the Pacific 10 Western University of Health Sciences 11 309 East Second Street 12 Pomona, California 91766 13 U.S.A. 14 [email protected] 15 16 17 Abstract 18 19 Objectives: As a primary trophic strategy, insectivory is uncommon and unevenly 20 distributed across extant primates. This pattern is partly a function of the challenges that 21 insectivory poses for large-bodied primates. In this study, I demonstrate that the uneven 22 distribution is also a consequence of variation in the rate of trophic evolution among small- 23 bodied lineages. 24 Methods: The sample consisted of 307 species classified by primary trophic strategy and 25 body size, creating an ordered three-state character: small-insectivorous, small-herbivorous, 26 and large-herbivorous. I tested for rate heterogeneity by partitioning major clades from the 27 rest of the primate tree and estimating separate rates of transition between herbivory and 28 insectivory for small-bodied lineages in each partition. 29 Results: Bayesian analysis of rate estimates indicates that a model with two rates of trophic 30 evolution provides the best fit to the data.