Living Mammals Are Placentals (Eutheria), Marsupials, and Monotremes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Miocene Paleotemperatures
QUARTERLY NEWSLETTER OF THE CALVERT MARINE MUSEUM FOSSIL CLUB Volume 3, Number 3 Editor: Sandy Roberts Whole Number 9 Summer 1987 MIOCENE PALEOTEMPERATURES By Reese Barrick University of Southern California studies of ocean paleotemperatures are important for increasing our knowledge of past oceans and climates. These studies have principally been carried out by using oxygen isotope ratios in fossils as paleother• mometers. The oxygen-18 to oxygen-16 (180/16/0) ratios incorporated into the shell and bone of marine animals reflect the isotopic composition and the temperature of the seawater in which they live. Most paleotemperature studies have used carbonate fossils (molluscs and certain microfossils) as the recorders of the ancient isotope history. This method however is complicated by several problems. The carbonate fossil may have had the original isotopic ratio which was incorporated into the shell while the animal was alive altered after it had died and undergone burial. These processes of alteration are represented by the term diagenesis. These diagenetic processes include overgrowth of the fossil by calcium carbonate in the sediment which may have different iso• topic ratios from the original fossil and would change the initial ratio. Another process which changes the initial isotopic ratio is the ready ex• change of oxygen atoms between seawater and carbonate atoms of the fossil. It has been shown by researchers that using phosphatic fossils as recorders of th~ oxygen isotope history of the ocean avoids the problems of diagenesis. The exchange of oxygen atoms between phosphate ions and seawater is negligible at low temperatures (lOO-2000C) unless biologically catalyzed during which exchange is extremely rapid. -

JVP 26(3) September 2006—ABSTRACTS
Neoceti Symposium, Saturday 8:45 acid-prepared osteolepiforms Medoevia and Gogonasus has offered strong support for BODY SIZE AND CRYPTIC TROPHIC SEPARATION OF GENERALIZED Jarvik’s interpretation, but Eusthenopteron itself has not been reexamined in detail. PIERCE-FEEDING CETACEANS: THE ROLE OF FEEDING DIVERSITY DUR- Uncertainty has persisted about the relationship between the large endoskeletal “fenestra ING THE RISE OF THE NEOCETI endochoanalis” and the apparently much smaller choana, and about the occlusion of upper ADAM, Peter, Univ. of California, Los Angeles, Los Angeles, CA; JETT, Kristin, Univ. of and lower jaw fangs relative to the choana. California, Davis, Davis, CA; OLSON, Joshua, Univ. of California, Los Angeles, Los A CT scan investigation of a large skull of Eusthenopteron, carried out in collaboration Angeles, CA with University of Texas and Parc de Miguasha, offers an opportunity to image and digital- Marine mammals with homodont dentition and relatively little specialization of the feeding ly “dissect” a complete three-dimensional snout region. We find that a choana is indeed apparatus are often categorized as generalist eaters of squid and fish. However, analyses of present, somewhat narrower but otherwise similar to that described by Jarvik. It does not many modern ecosystems reveal the importance of body size in determining trophic parti- receive the anterior coronoid fang, which bites mesial to the edge of the dermopalatine and tioning and diversity among predators. We established relationships between body sizes of is received by a pit in that bone. The fenestra endochoanalis is partly floored by the vomer extant cetaceans and their prey in order to infer prey size and potential trophic separation of and the dermopalatine, restricting the choana to the lateral part of the fenestra. -

PDF of Manuscript and Figures
The triple origin of whales DAVID PETERS Independent Researcher 311 Collinsville Avenue, Collinsville, IL 62234, USA [email protected] 314-323-7776 July 13, 2018 RH: PETERS—TRIPLE ORIGIN OF WHALES Keywords: Cetacea, Mysticeti, Odontoceti, Phylogenetic analyis ABSTRACT—Workers presume the traditional whale clade, Cetacea, is monophyletic when they support a hypothesis of relationships for baleen whales (Mysticeti) rooted on stem members of the toothed whale clade (Odontoceti). Here a wider gamut phylogenetic analysis recovers Archaeoceti + Odontoceti far apart from Mysticeti and right whales apart from other mysticetes. The three whale clades had semi-aquatic ancestors with four limbs. The clade Odontoceti arises from a lineage that includes archaeocetids, pakicetids, tenrecs, elephant shrews and anagalids: all predators. The clade Mysticeti arises from a lineage that includes desmostylians, anthracobunids, cambaytheres, hippos and mesonychids: none predators. Right whales are derived from a sister to Desmostylus. Other mysticetes arise from a sister to the RBCM specimen attributed to Behemotops. Basal mysticetes include Caperea (for right whales) and Miocaperea (for all other mysticetes). Cetotheres are not related to aetiocetids. Whales and hippos are not related to artiodactyls. Rather the artiodactyl-type ankle found in basal archaeocetes is also found in the tenrec/odontocete clade. Former mesonychids, Sinonyx and Andrewsarchus, nest close to tenrecs. These are novel observations and hypotheses of mammal interrelationships based on morphology and a wide gamut taxon list that includes relevant taxa that prior studies ignored. Here some taxa are tested together for the first time, so they nest together for the first time. INTRODUCTION Marx and Fordyce (2015) reported the genesis of the baleen whale clade (Mysticeti) extended back to Zygorhiza, Physeter and other toothed whales (Archaeoceti + Odontoceti). -

Mandible Allometry in Extant and Fossil Balaenopteridae (Cetacea: Mammalia): the Largest Vertebrate Skeletal Element and Its Role in Rorqual Lunge Feeding
bs_bs_banner Biological Journal of the Linnean Society, 2013, 108, 586–599. With 6 figures Mandible allometry in extant and fossil Balaenopteridae (Cetacea: Mammalia): the largest vertebrate skeletal element and its role in rorqual lunge feeding NICHOLAS D. PYENSON1,2*, JEREMY A. GOLDBOGEN3 and ROBERT E. SHADWICK4 1Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, P.O. Box 37012, Washington, DC, 20013-7013, USA 2Departments of Mammalogy and Paleontology, Burke Museum of Natural History and Culture, Seattle, WA 98195, USA 3Cascadia Research Collective, 218½ West 4th Avenue, Olympia, WA, USA 4Department of Zoology, University of British Columbia, 6270 University Boulevard, Vancouver, BC, Canada V6T 1Z4 Received 4 July 2012; revised 10 September 2012; accepted for publication 10 September 2012 Rorqual whales (crown Balaenopteridae) are unique among aquatic vertebrates in their ability to lunge feed. During a single lunge, rorquals rapidly engulf a large volume of prey-laden water at high speed, which they then filter to capture suspended prey. Engulfment biomechanics are mostly governed by the coordinated opening and closing of the mandibles at large gape angles, which differentially exposes the floor of the oral cavity to oncoming flow. The mouth area in rorquals is delimited by unfused bony mandibles that form kinetic linkages to each other and with the skull. The relative scale and morphology of these skeletal elements have profound consequences for the energetic efficiency of foraging in these gigantic predators. Here, we performed a morphometric study of rorqual mandibles using a data set derived from a survey of museum specimens. Across adult specimens of extant balaenopterids, mandibles range in size from ~1–6 m in length, and at their upper limit they represent the single largest osteological element of any vertebrate, living or extinct. -

8. Primate Evolution
8. Primate Evolution Jonathan M. G. Perry, Ph.D., The Johns Hopkins University School of Medicine Stephanie L. Canington, B.A., The Johns Hopkins University School of Medicine Learning Objectives • Understand the major trends in primate evolution from the origin of primates to the origin of our own species • Learn about primate adaptations and how they characterize major primate groups • Discuss the kinds of evidence that anthropologists use to find out how extinct primates are related to each other and to living primates • Recognize how the changing geography and climate of Earth have influenced where and when primates have thrived or gone extinct The first fifty million years of primate evolution was a series of adaptive radiations leading to the diversification of the earliest lemurs, monkeys, and apes. The primate story begins in the canopy and understory of conifer-dominated forests, with our small, furtive ancestors subsisting at night, beneath the notice of day-active dinosaurs. From the archaic plesiadapiforms (archaic primates) to the earliest groups of true primates (euprimates), the origin of our own order is characterized by the struggle for new food sources and microhabitats in the arboreal setting. Climate change forced major extinctions as the northern continents became increasingly dry, cold, and seasonal and as tropical rainforests gave way to deciduous forests, woodlands, and eventually grasslands. Lemurs, lorises, and tarsiers—once diverse groups containing many species—became rare, except for lemurs in Madagascar where there were no anthropoid competitors and perhaps few predators. Meanwhile, anthropoids (monkeys and apes) emerged in the Old World, then dispersed across parts of the northern hemisphere, Africa, and ultimately South America. -

Early Eocene Primates from Gujarat, India
ARTICLE IN PRESS Journal of Human Evolution xxx (2009) 1–39 Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Early Eocene Primates from Gujarat, India Kenneth D. Rose a,*, Rajendra S. Rana b, Ashok Sahni c, Kishor Kumar d, Pieter Missiaen e, Lachham Singh b, Thierry Smith f a Johns Hopkins University School of Medicine, Baltimore, Maryland 21205, USA b H.N.B. Garhwal University, Srinagar 246175, Uttarakhand, India c Panjab University, Chandigarh 160014, India d Wadia Institute of Himalayan Geology, Dehradun 248001, Uttarakhand, India e University of Ghent, B-9000 Ghent, Belgium f Royal Belgian Institute of Natural Sciences, B-1000 Brussels, Belgium article info abstract Article history: The oldest euprimates known from India come from the Early Eocene Cambay Formation at Vastan Mine Received 24 June 2008 in Gujarat. An Ypresian (early Cuisian) age of w53 Ma (based on foraminifera) indicates that these Accepted 8 January 2009 primates were roughly contemporary with, or perhaps predated, the India-Asia collision. Here we present new euprimate fossils from Vastan Mine, including teeth, jaws, and referred postcrania of the Keywords: adapoids Marcgodinotius indicus and Asiadapis cambayensis. They are placed in the new subfamily Eocene Asiadapinae (family Notharctidae), which is most similar to primitive European Cercamoniinae such as India Donrussellia and Protoadapis. Asiadapines were small primates in the size range of extant smaller Notharctidae Adapoidea bushbabies. Despite their generally very plesiomorphic morphology, asiadapines also share a few derived Omomyidae dental traits with sivaladapids, suggesting a possible relationship to these endemic Asian adapoids. In Eosimiidae addition to the adapoids, a new species of the omomyid Vastanomys is described. -

The Taxonomic and Evolutionary History of Fossil and Modern Balaenopteroid Mysticetes
Journal of Mammalian Evolution, Vol. 12, Nos. 1/2, June 2005 (C 2005) DOI: 10.1007/s10914-005-6944-3 The Taxonomic and Evolutionary History of Fossil and Modern Balaenopteroid Mysticetes Thomas A. Demer´ e,´ 1,4 Annalisa Berta,2 and Michael R. McGowen2,3 Balaenopteroids (Balaenopteridae + Eschrichtiidae) are a diverse lineage of living mysticetes, with seven to ten species divided between three genera (Megaptera, Balaenoptera and Eschrichtius). Extant members of the Balaenopteridae (Balaenoptera and Megaptera) are characterized by their engulfment feeding behavior, which is associated with a number of unique cranial, mandibular, and soft anatomical characters. The Eschrichtiidae employ suction feeding, which is associated with arched rostra and short, coarse baleen. The recognition of these and other characters in fossil balaenopteroids, when viewed in a phylogenetic framework, provides a means for assessing the evolutionary history of this clade, including its origin and diversification. The earliest fossil balaenopterids include incomplete crania from the early late Miocene (7–10 Ma) of the North Pacific Ocean Basin. Our preliminary phylogenetic results indicate that the basal taxon, “Megaptera” miocaena should be reassigned to a new genus based on its possession of primitive and derived characters. The late late Miocene (5–7 Ma) balaenopterid record, except for Parabalaenoptera baulinensis and Balaenoptera siberi, is largely undescribed and consists of fossil specimens from the North and South Pacific and North Atlantic Ocean basins. The Pliocene record (2–5 Ma) is very diverse and consists of numerous named, but problematic, taxa from Italy and Belgium, as well as unnamed taxa from the North and South Pacific and eastern North Atlantic Ocean basins. -

The Biology of Marine Mammals
Romero, A. 2009. The Biology of Marine Mammals. The Biology of Marine Mammals Aldemaro Romero, Ph.D. Arkansas State University Jonesboro, AR 2009 2 INTRODUCTION Dear students, 3 Chapter 1 Introduction to Marine Mammals 1.1. Overture Humans have always been fascinated with marine mammals. These creatures have been the basis of mythical tales since Antiquity. For centuries naturalists classified them as fish. Today they are symbols of the environmental movement as well as the source of heated controversies: whether we are dealing with the clubbing pub seals in the Arctic or whaling by industrialized nations, marine mammals continue to be a hot issue in science, politics, economics, and ethics. But if we want to better understand these issues, we need to learn more about marine mammal biology. The problem is that, despite increased research efforts, only in the last two decades we have made significant progress in learning about these creatures. And yet, that knowledge is largely limited to a handful of species because they are either relatively easy to observe in nature or because they can be studied in captivity. Still, because of television documentaries, ‘coffee-table’ books, displays in many aquaria around the world, and a growing whale and dolphin watching industry, people believe that they have a certain familiarity with many species of marine mammals (for more on the relationship between humans and marine mammals such as whales, see Ellis 1991, Forestell 2002). As late as 2002, a new species of beaked whale was being reported (Delbout et al. 2002), in 2003 a new species of baleen whale was described (Wada et al. -

Evidence for an Asian Origin of Stem Anthropoids
Evidence for an Asian origin of stem anthropoids Richard F. Kay1 Department of Evolutionary Anthropology, Duke University, Durham, NC 27708-03083 n PNAS, Chaimanee et al. (1) report Genetic, embryological, and anatomical We are not there yet. Recent comprehen- a previously undescribed species of evidence demonstrates that the sister sive phylogenetic analyses stemming from I primate, Afrasia, from the late Middle group of Anthropoidea is the south Asian virtually the same datasets yield somewhat Eocene of Burma. They identify tarsier, with the two forming the crown different cladograms that reflect sensitivity Afrasia as the sister taxon to the African group Haplorhini (6–8). The other clade of to which taxa are included in the analysis genus Afrotarsius but slightly more primitive extant primates is the lemurs and lorises, and which sets of analytical assumptions are than it and allied with stem Anthropoidea called Strepsirrhini. Eocene Holarctic selected. Pertinent to the biogeographic of south Asia. Anthropoidea is the taxo- Omomyoidea are generally considered as conclusions of Chaimanee et al. (1), Seiffert nomic group that today includes New and stem haplorhines, although the precise et al. (11) conclude that Afrotarsius cha- Old World monkeys, apes, and humans. If relationship of omomyoids to tarsiers and trathi [a younger species than the one that upheld, the biogeographic significance of anthropoids is uncertain (8). Tarsius often Chaimanee et al. (1) describe] is an African these results is profound: If Afrasia and is considered to be a relictual omomyoid tarsioid. If Seiffert et al. (11) are correct, Afrotarsius are as closely related as Chai- in south Asia. -
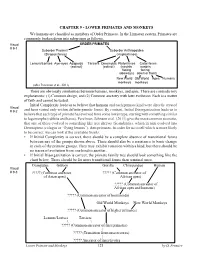
CHAPTER 9 - LOWER PRIMATES and MONKEYS We Humans Are Classified As Members of Order Primates
CHAPTER 9 - LOWER PRIMATES AND MONKEYS We humans are classified as members of Order Primates. In the Linnaean system, Primates are commonly broken down into subgroups as follows. Visual ORDER PRIMATES # 9-1 Suborder Prosimii Suborder Anthropoidea (Strepsirrhines) (Haplorrhines) Lemurs/Lorises Aye-ayes Adapoids Tarsiers Omomyids Platyrrhines Catarrhines (extinct) (extinct) (nostrils nostrils facing facing sideways) down or front) New World Old World Apes Humans monkeys monkeys (after Perelman et al., 2011) There are obviously similarities between humans, monkeys, and apes. There are contradictory explanations: (1) Common design, and (2) Common ancestry with later evolution. Each is a matter of faith and cannot be tested. Initial Complexity leads us to believe that humans and each primate kind were directly created Visual # 9-2 and have varied only within definite genetic limits. By contrast, Initial Disorganization leads us to believe that each type of primate has evolved from some lower type, starting with something similar to lagomorphs (rabbits and hares). Perelman, Johnson et al. (2011) give the most common scenario, that one of these evolved to something like tree shrews (Scandentia), which in turn evolved into Dermoptera (colugos or “flying lemurs”), then primates. In order for us to tell which is more likely to be correct, we can look at the available fossils. • If Initial Complexity is correct, there should be a complete absence of transitional forms between any of the groups shown above. There should also be a resistance to basic change in each of the primate groups. They may exhibit variation within a kind, but there should be no traces of evolution from one kind to another. -

The Tempo of Trophic Evolution in Small-Bodied Primates 2 3 4 Running Title: Trophic Evolution in Small-Bodied Primates 5 6 7 Jeremiah E
bioRxiv preprint doi: https://doi.org/10.1101/2020.03.17.996207; this version posted March 20, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-ND 4.0 International license. 1 1 The tempo of trophic evolution in small-bodied primates 2 3 4 Running title: Trophic evolution in small-bodied primates 5 6 7 Jeremiah E. Scott 8 Department of Medical Anatomical Sciences 9 College of Osteopathic Medicine of the Pacific 10 Western University of Health Sciences 11 309 East Second Street 12 Pomona, California 91766 13 U.S.A. 14 [email protected] 15 16 17 Abstract 18 19 Objectives: As a primary trophic strategy, insectivory is uncommon and unevenly 20 distributed across extant primates. This pattern is partly a function of the challenges that 21 insectivory poses for large-bodied primates. In this study, I demonstrate that the uneven 22 distribution is also a consequence of variation in the rate of trophic evolution among small- 23 bodied lineages. 24 Methods: The sample consisted of 307 species classified by primary trophic strategy and 25 body size, creating an ordered three-state character: small-insectivorous, small-herbivorous, 26 and large-herbivorous. I tested for rate heterogeneity by partitioning major clades from the 27 rest of the primate tree and estimating separate rates of transition between herbivory and 28 insectivory for small-bodied lineages in each partition. 29 Results: Bayesian analysis of rate estimates indicates that a model with two rates of trophic 30 evolution provides the best fit to the data. -

How Modern Baleen Whales Arose from a Fossil “Dark Age”
Editors' choice Like phoenix from the ashes: How modern baleen whales arose from a fossil “dark age” FELIX G. MARX, ERICH M.G. FITZGERALD, and R. EWAN FORDYCE Marx, F.G., Fitzgerald, E.M.G., and Fordyce, R.E. 2019. Like phoenix from the ashes: How modern baleen whales arose from a fossil “dark age”. Acta Palaeontologica Polonica 64 (2): 231–238. The evolution of baleen whales (Mysticeti), the largest animals on Earth, was punctuated by a pivotal turnover event. Following their emergence around 36 million years (Ma), mysticetes diversified into a disparate range of toothed and toothless species until 23 Ma, but then nearly vanished from the global fossil record for the next five million years. Following this early Miocene “dark age”, toothless mysticetes spectacularly reappeared around 18–17 Ma, whereas toothed mysticetes had gone entirely extinct. Here, we suggest that this turnover event reflects a change in mysticete habitat occupancy. Using the well-sampled record of Australasia as a case study, we show that Oligocene pre-“dark age” mysticetes formed distinct coastal and offshore assemblages, dominated by small (2–4 m), ecologically disparate toothed species, and larger (5–6 m) toothless filter feeders, respectively. Environmental change around the Oligocene–Miocene boundary led to the decline of the endemic coastal assemblages, leaving nearshore deposits virtually devoid of mys- ticetes. Filter feeders persisted offshore and subsequently re-invaded coastal habitats during the mid-Miocene Climatic Optimum, thus establishing the modern, cosmopolitan mysticete fauna. Key words: Mammalia, Mysticeti, evolution, Oligocene, Miocene, Zealandia, Australia. Felix G. Marx [[email protected]], Directorate Earth and History of Life, Royal Belgian Institute of Natural Sciences, Rue Vautier 29, Brussels 1000, Belgium; Department of Geology, University of Liège, Belgium; School of Biological Sciences, Monash University, Clayton, Vic., Australia; Museums Victoria, Melbourne, Vic., Australia.