A New Clade of Basal Early Cretaceous Pygostylian Birds and Developmental Plasticity of the Avian Shoulder Girdle
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Reexamination of Four Prolacertiforms \Tith Implications for Pterosaur Phylogenesis
Rìvista Italiana di Paleontologia e Stratigrafia Dicembre 2000 I--r4-""l*-I-."-''* 1 A REEXAMINATION OF FOUR PROLACERTIFORMS \TITH IMPLICATIONS FOR PTEROSAUR PHYLOGENESIS DAVID PETERS ReceìterJ October 23, 1999; accepted October 20, 200A Kqt uorcls: Pterosauria, Prolacertiiormes (Reprilia, Diapsida), Traditionally the answer has been rhat prerosaurs Phyìogeny, Cladisric an:ìy.is. are archosaurs (Romer 1956); the sister group of the Riassunto . Tradizionalmente gli prerosauri venir.ano considerati Dinosauria, ScleromochÌus a.nd Lagosuclcws/Maraswchus come appartenenti agli Archosaurifomes e molti specìalistì contempo_ (Benton 1985, 1990, 1999; Padian 1984; Gauthter 1984, ranei considerano gli pterosauri quali sisrer groups di Lagosuchus, 1986; Sereno 1991, 1994; Kellner 1996); or perhaps Schleromochlus e dei Dinosauria. La nuova analisi filogenerica qui pro- archosauriformes close posta merte in discussione queste affinirà jn quanto tutte le presunte to prorerosuchids and eryrhro- sinapomorfie che collegherebbero gli Pterosauria con gli Archosauri_ suchids (Bennett 1996a), chiefly because prerosaurs formes o con gli pterosaurìa, Ornìthodira mancano in realtà negli have a prominent anrorbiral fenestra and a suite of other oppure sono condivise anche da alcuni taxa di prolacertiformi. ll archosaur-like characrers almosr entirely recente riesame degli olotipi dt confined to the Cosesaurus a,Liceps, Longisquama ìnsig_ hind nis e di Sharovipteryx mìrabi/ìs suggeriscono che molti caratteri potreb- limb (Bennert 1996a). Although Benton (1982, bero venire interpretati in maniera diversa rispetto alle precedenti L984) initially indicated that the prerosauria are descrìzioni. I risultati di molteplici analisì cladistjche suggeriscono che archosauromorphs and the sister-group ro all other questi tre prolacertìformi enigmatici, uniramente a Langobardìsawrws, archosauromorphs, later work (Benton 1985, 1.990, recentemente descritto, costituirebbero i sister taxa degli prerosauri, in base ad un insieme di sinapomorfie di nuova identificazione. -

A New Clade of Basal Early Cretaceous Pygostylian Birds and Developmental Plasticity of the Avian Shoulder Girdle
A new clade of basal Early Cretaceous pygostylian birds and developmental plasticity of the avian shoulder girdle Min Wanga,b,1, Thomas A. Stidhama,b, and Zhonghe Zhoua,b,1 aKey Laboratory of Vertebrate Evolution and Human Origins, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China; and bCenter for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Beijing 100044, China Contributed by Zhonghe Zhou, August 16, 2018 (sent for review July 16, 2018; reviewed by Stephen L. Brusatte and Gareth Dyke) Early members of the clade Pygostylia (birds with a short tail Systematic Paleontology ending in a compound bone termed “pygostyle”)arecriticalfor Aves Linnaeus, 1758; Pygostylia Chiappe, 2002; Jinguofortisidae understanding how the modern avian bauplan evolved from fam. nov. (SI Appendix, SI Text); Jinguofortis perplexus gen. et sp. nov. long-tailed basal birds like Archaeopteryx. However, the cur- rently limited known diversity of early branching pygostylians Holotype obscures our understanding of this major transition in avian A complete and articulated skeleton with feathers is housed at evolution. Here, we describe a basal pygostylian, Jinguofortis the Institute of Vertebrate Paleontology and Paleoanthropology perplexus gen. et sp. nov., from the Early Cretaceous of China (IVPP) under the collect number IVPP V24194 (Fig. 1 and SI that adds important information about early members of the Appendix, Figs. S1–S7 and Table S1). short-tailed bird group. Phylogenetic analysis recovers a clade (Jinguofortisidae fam. nov.) uniting Jinguofortis and the enig- Etymology matic basal avian taxon Chongmingia that represents the second The generic name is derived from “jinguo” (Mandarin), referring earliest diverging group of the Pygostylia. -

New Oviraptorid Dinosaur (Dinosauria: Oviraptorosauria) from the Nemegt Formation of Southwestern Mongolia
Bull. Natn. Sci. Mus., Tokyo, Ser. C, 30, pp. 95–130, December 22, 2004 New Oviraptorid Dinosaur (Dinosauria: Oviraptorosauria) from the Nemegt Formation of Southwestern Mongolia Junchang Lü1, Yukimitsu Tomida2, Yoichi Azuma3, Zhiming Dong4 and Yuong-Nam Lee5 1 Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China 2 National Science Museum, 3–23–1 Hyakunincho, Shinjukuku, Tokyo 169–0073, Japan 3 Fukui Prefectural Dinosaur Museum, 51–11 Terao, Muroko, Katsuyama 911–8601, Japan 4 Institute of Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China 5 Korea Institute of Geoscience and Mineral Resources, Geology & Geoinformation Division, 30 Gajeong-dong, Yuseong-gu, Daejeon 305–350, South Korea Abstract Nemegtia barsboldi gen. et sp. nov. here described is a new oviraptorid dinosaur from the Late Cretaceous (mid-Maastrichtian) Nemegt Formation of southwestern Mongolia. It differs from other oviraptorids in the skull having a well-developed crest, the anterior margin of which is nearly vertical, and the dorsal margin of the skull and the anterior margin of the crest form nearly 90°; the nasal process of the premaxilla being less exposed on the dorsal surface of the skull than those in other known oviraptorids; the length of the frontal being approximately one fourth that of the parietal along the midline of the skull. Phylogenetic analysis shows that Nemegtia barsboldi is more closely related to Citipati osmolskae than to any other oviraptorosaurs. Key words : Nemegt Basin, Mongolia, Nemegt Formation, Late Cretaceous, Oviraptorosauria, Nemegtia. dae, and Caudipterygidae (Barsbold, 1976; Stern- Introduction berg, 1940; Currie, 2000; Clark et al., 2001; Ji et Oviraptorosaurs are generally regarded as non- al., 1998; Zhou and Wang, 2000; Zhou et al., avian theropod dinosaurs (Osborn, 1924; Bars- 2000). -

Appendix A. Supplementary Material
Appendix A. Supplementary material Comprehensive taxon sampling and vetted fossils help clarify the time tree of shorebirds (Aves, Charadriiformes) David Cernˇ y´ 1,* & Rossy Natale2 1Department of the Geophysical Sciences, University of Chicago, Chicago 60637, USA 2Department of Organismal Biology & Anatomy, University of Chicago, Chicago 60637, USA *Corresponding Author. Email: [email protected] Contents 1 Fossil Calibrations 2 1.1 Calibrations used . .2 1.2 Rejected calibrations . 22 2 Outgroup sequences 30 2.1 Neornithine outgroups . 33 2.2 Non-neornithine outgroups . 39 3 Supplementary Methods 72 4 Supplementary Figures and Tables 74 5 Image Credits 91 References 99 1 1 Fossil Calibrations 1.1 Calibrations used Calibration 1 Node calibrated. MRCA of Uria aalge and Uria lomvia. Fossil taxon. Uria lomvia (Linnaeus, 1758). Specimen. CASG 71892 (referred specimen; Olson, 2013), California Academy of Sciences, San Francisco, CA, USA. Lower bound. 2.58 Ma. Phylogenetic justification. As in Smith (2015). Age justification. The status of CASG 71892 as the oldest known record of either of the two spp. of Uria was recently confirmed by the review of Watanabe et al. (2016). The younger of the two marine transgressions at the Tolstoi Point corresponds to the Bigbendian transgression (Olson, 2013), which contains the Gauss-Matuyama magnetostratigraphic boundary (Kaufman and Brigham-Grette, 1993). Attempts to date this reversal have been recently reviewed by Ohno et al. (2012); Singer (2014), and Head (2019). In particular, Deino et al. (2006) were able to tightly bracket the age of the reversal using high-precision 40Ar/39Ar dating of two tuffs in normally and reversely magnetized lacustrine sediments from Kenya, obtaining a value of 2.589 ± 0.003 Ma. -
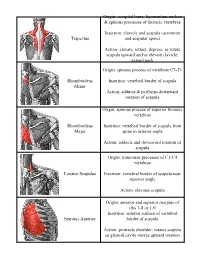
Trapezius Origin: Occipital Bone, Ligamentum Nuchae & Spinous Processes of Thoracic Vertebrae Insertion: Clavicle and Scapul
Origin: occipital bone, ligamentum nuchae & spinous processes of thoracic vertebrae Insertion: clavicle and scapula (acromion Trapezius and scapular spine) Action: elevate, retract, depress, or rotate scapula upward and/or elevate clavicle; extend neck Origin: spinous process of vertebrae C7-T1 Rhomboideus Insertion: vertebral border of scapula Minor Action: adducts & performs downward rotation of scapula Origin: spinous process of superior thoracic vertebrae Rhomboideus Insertion: vertebral border of scapula from Major spine to inferior angle Action: adducts and downward rotation of scapula Origin: transverse precesses of C1-C4 vertebrae Levator Scapulae Insertion: vertebral border of scapula near superior angle Action: elevates scapula Origin: anterior and superior margins of ribs 1-8 or 1-9 Insertion: anterior surface of vertebral Serratus Anterior border of scapula Action: protracts shoulder: rotates scapula so glenoid cavity moves upward rotation Origin: anterior surfaces and superior margins of ribs 3-5 Insertion: coracoid process of scapula Pectoralis Minor Action: depresses & protracts shoulder, rotates scapula (glenoid cavity rotates downward), elevates ribs Origin: supraspinous fossa of scapula Supraspinatus Insertion: greater tuberacle of humerus Action: abduction at the shoulder Origin: infraspinous fossa of scapula Infraspinatus Insertion: greater tubercle of humerus Action: lateral rotation at shoulder Origin: clavicle and scapula (acromion and adjacent scapular spine) Insertion: deltoid tuberosity of humerus Deltoid Action: -

The Scapulocoracoid of an Early Triassic Stem−Frog from Poland
The scapulocoracoid of an Early Triassic stem−frog from Poland MAGDALENA BORSUK−BIAŁYNICKA and SUSAN E. EVANS Borsuk−Białynicka, M. and Evans, S.E. 2002. The scapulocoracoid of an Early Triassic stem−frog from Poland. Acta Palaeontologica Polonica 47 (1): 79–96. The scapulocoracoid of Czatkobatrachus polonicus Evans and Borsuk−Białynicka, 1998, a stem−frog from the Early Tri− assic karst locality of Czatkowice (Southern Poland), is described. The overall type of scapulocoracoid is plesiomorphic, but the subcircular shape and laterally oriented glenoid is considered synapomorphic of Salientia. The supraglenoid fora− men is considered homologous to the scapular cleft of the Anura. In Czatkobatrachus, the supraglenoid foramen occupies an intermediate position between that of the early tetrapod foramen and the scapular cleft of Anura. The cleft scapula is probably synapomorphic for the Anura. In early salientian phylogeny, the shift in position of the supraglenoid foramen may have been associated with an anterior rotation of the forelimb. This change in position of the forelimb may reflect an evolutionary shift from a mainly locomotory function to static functions (support, balance, eventually shock−absorption). Laterally extended limbs may have been more effective than posterolateral ones in absorbing landing stresses, until the specialised shock−absorption pectoral mechanism of crown−group Anura had developed. The glenoid shape and position, and the slender scapular blade, of Czatkobatrachus, in combination with the well−ossified joint surfaces on the humerus and ulna, all support a primarily terrestrial rather than aquatic mode of life. The new Polish material also permits clarifica− tion of the pectoral anatomy of the contemporaneous Madagascan genus Triadobatrachus. -

Reidentification of Avian Embryonic Remains from the Cretaceous of Mongolia
RESEARCH ARTICLE Reidentification of Avian Embryonic Remains from the Cretaceous of Mongolia David J. Varricchio1*, Amy M. Balanoff2, Mark A. Norell3 1 Department of Earth Sciences, Montana State University, Bozeman, Montana, 59717, United States of America, 2 Department of Anatomical Sciences, Stony Brook University School of Medicine, Stony Brook, NY, 11794, United States of America, 3 Division of Paleontology, American Museum of Natural History, New York, NY, 10024, United States of America * [email protected] Abstract Embryonic remains within a small (4.75 by 2.23 cm) egg from the Late Cretaceous, Mongo- lia are here re-described. High-resolution X-ray computed tomography (HRCT) was used to digitally prepare and describe the enclosed embryonic bones. The egg, IGM (Mongolian In- stitute for Geology, Ulaanbaatar) 100/2010, with a three-part shell microstructure, was originally assigned to Neoceratopsia implying extensive homoplasy among eggshell char- acters across Dinosauria. Re-examination finds the forelimb significantly longer than the hindlimbs, proportions suggesting an avian identification. Additional, postcranial apomor- phies (strut-like coracoid, cranially located humeral condyles, olecranon fossa, slender radi- OPEN ACCESS us relative to the ulna, trochanteric crest on the femur, and ulna longer than the humerus) identify the embryo as avian. Presence of a dorsal coracoid fossa and a craniocaudally Citation: Varricchio DJ, Balanoff AM, Norell MA (2015) Reidentification of Avian Embryonic Remains compressed distal humerus with a strongly angled distal margin support a diagnosis of IGM from the Cretaceous of Mongolia. PLoS ONE 10(6): 100/2010 as an enantiornithine. Re-identification eliminates the implied homoplasy of this e0128458. doi:10.1371/journal.pone.0128458 tri-laminate eggshell structure, and instead associates enantiornithine birds with eggshell Academic Editor: Peter Dodson, University of microstructure composed of a mammillary, squamatic, and external zones. -

Coracoid Process Anatomy: a Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B
Quantitative Assessment of the Coracoacromial and the Coracoclavicular Ligaments With 3-Dimensional Mapping of the Coracoid Process Anatomy: A Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B. Ferrari, M.D., George Sanchez, B.S., Alex W. Brady, M.Sc., Jonas Pogorzelski, M.D., M.H.B.A., George F. Lebus, M.D., Peter J. Millett, M.D., M.Sc., Robert F. LaPrade, M.D., Ph.D., and CAPT Matthew T. Provencher, M.D., M.C., U.S.N.R. Purpose: To perform a quantitative anatomic evaluation of the (1) coracoid process, specifically the attachment sites of the conjoint tendon, the pectoralis minor, the coracoacromial ligament (CAL), and the coracoclavicular (CC) ligaments in relation to pertinent osseous and soft tissue landmarks; (2) CC ligaments’ attachments on the clavicle; and (3) CAL attachment on the acromion in relation to surgically relevant anatomic landmarks to assist in planning of the Latarjet procedure, acromioclavicular (AC) joint reconstructions, and CAL resection distances avoiding iatrogenic injury to sur- rounding structures. Methods: Ten nonpaired fresh-frozen human cadaveric shoulders (mean age 52 years, range 33- 64 years) were included in this study. A 3-dimensional coordinate measuring device was used to quantify the location of pertinent bony landmarks and soft tissue attachment areas. The ligament and tendon attachment perimeters and center points on the coracoid, clavicle, and acromion were identified and subsequently dissected off the bone. Coordinates of points along the perimeters of attachment sites were used to calculate areas, whereas coordinates of center points were used to determine distances between surgically relevant attachment sites and pertinent bony landmarks. -

Featured Article Cranial Anatomy of Erlikosaurus Andrewsi (Dinosauria, Therizinosauria): New Insights Based on Digital Reconstru
Journal of Vertebrate Paleontology 34(6):1263–1291, November 2014 Ó 2014 by the Society of Vertebrate Paleontology FEATURED ARTICLE CRANIAL ANATOMY OF ERLIKOSAURUS ANDREWSI (DINOSAURIA, THERIZINOSAURIA): NEW INSIGHTS BASED ON DIGITAL RECONSTRUCTION STEPHAN LAUTENSCHLAGER,*,1 LAWRENCE M. WITMER,2 PERLE ALTANGEREL,3 LINDSAY E. ZANNO,4,5 and EMILY J. RAYFIELD1 1School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K., [email protected]; 2Department of Biomedical Sciences, Heritage College of Osteopathic Medicine, Ohio University, Athens, Ohio 45701, U.S.A.; 3National University of Mongolia, Ulaanbaatar, Mongolia; 4Nature Research Center, NC Museum of Natural Sciences, Raleigh, North Carolina 27695, U.S.A.; 5Department of Biology, North Carolina State University, Raleigh, North Carolina 27601, U.S.A. ABSTRACT—The skull of Erlikosaurus andrewsi from the Upper Cretaceous Baishin Tsav locality of Mongolia represents the only known three-dimensionally preserved and nearly complete skull of a therizinosaurian. Computed tomographic (CT) scanning of the original specimen and three-dimensional visualization techniques allow the cranial skeleton to be digitally prepared, disarticulated, and restored. Here, we present a detailed description of the restored skull morphology and the individual cranial elements, including visualization of the internal neurovascular and pneumatic structures. Information gained from this study is used in a revised and emended diagnosis for E. andrewsi. A reappraisal of the evolutionary and functional changes in the cranial skeleton as provided by this study supports prior proposals that a keratinous sheath or rhamphotheca was developed early in the evolution of Therizinosauria. Paralleled by the reduction of functional and replacement teeth, this development indicates a shift in the manner of food processing/procurement at the tip of the snout. -

Reconstructing Pectoral Appendicular Muscle Anatomy in Fossil Fish and Tetrapods Over the Fins-To-Limbs Transition
Biol. Rev. (2017), pp. 000–000. 1 doi: 10.1111/brv.12386 Reconstructing pectoral appendicular muscle anatomy in fossil fish and tetrapods over the fins-to-limbs transition Julia L. Molnar1,∗ , Rui Diogo2, John R. Hutchinson3 and Stephanie E. Pierce4 1Department of Anatomy, New York Institute of Technology College of Osteopathic Medicine, Northern Boulevard, Old Westbury, NY, U.S.A. 2Department of Anatomy, Howard University College of Medicine, 520 W St. NW, Numa Adams Building, Washington, DC 20059, U.S.A. 3Structure and Motion Lab, Royal Veterinary College, Hawkshead Lane, Hatfield, Hertfordshire AL9 7TA, UK 4Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, U.S.A. ABSTRACT The question of how tetrapod limbs evolved from fins is one of the great puzzles of evolutionary biology. While palaeontologists, developmental biologists, and geneticists have made great strides in explaining the origin and early evolution of limb skeletal structures, that of the muscles remains largely unknown. The main reason is the lack of consensus about appendicular muscle homology between the closest living relatives of early tetrapods: lobe-finned fish and crown tetrapods. In the light of a recent study of these homologies, we re-examined osteological correlates of muscle attachment in the pectoral girdle, humerus, radius, and ulna of early tetrapods and their close relatives. Twenty-nine extinct and six extant sarcopterygians were included in a meta-analysis using information from the literature and from original specimens, when possible. We analysed these osteological correlates using parsimony-based character optimization in order to reconstruct muscle anatomy in ancestral lobe-finned fish, tetrapodomorph fish, stem tetrapods, and crown tetrapods. -

Breathing and Locomotion in Birds
Breathing and locomotion in birds. A thesis submitted to the University of Manchester for the degree of Doctor of Philosophy in the Faculty of Life Sciences. 2010 Peter George Tickle Contents Abstract 4 Declaration 5 Copyright Statement 6 Author Information 7 Acknowledgements 9 Organisation of this PhD thesis 10 Chapter 1 General Introduction 13 1. Introduction 14 1.1 The Avian Respiratory System 14 1.1.1 Structure of the lung and air sacs 16 1.1.2 Airflow in the avian respiratory system 21 1.1.3 The avian aspiration pump 25 1.2 The uncinate processes in birds 29 1.2.1 Uncinate process morphology and biomechanics 32 1.3 Constraints on breathing in birds 33 1.3.1 Development 33 1.3.2 Locomotion 35 1.3.2.1 The appendicular skeleton 35 1.3.2.2 Overcoming the trade-off between breathing 36 and locomotion 1.3.2.3 Energetics of locomotion in birds 38 1.4 Evolution of the ventilatory pump in birds 41 1.5 Overview and Thesis Aims 42 2 Chapter 2 Functional significance of the uncinate processes in birds. 44 Chapter 3 Ontogenetic development of the uncinate processes in the 45 domestic turkey (Meleagris gallopavo). Chapter 4 Uncinate process length in birds scales with resting metabolic rate. 46 Chapter 5 Load carrying during locomotion in the barnacle goose (Branta 47 leucopsis): The effect of load placement and size. Chapter 6 A continuum in ventilatory mechanics from early theropods to 48 extant birds. Chapter 7 General Discussion 49 References 64 3 Abstract of a thesis by Peter George Tickle submitted to the University of Manchester for the degree of PhD in the Faculty of Life Sciences and entitled ‘Breathing and Locomotion in Birds’. -

The Oldest Record of Ornithuromorpha from the Early Cretaceous of China
ARTICLE Received 6 Jan 2015 | Accepted 20 Mar 2015 | Published 5 May 2015 DOI: 10.1038/ncomms7987 OPEN The oldest record of ornithuromorpha from the early cretaceous of China Min Wang1, Xiaoting Zheng2,3, Jingmai K. O’Connor1, Graeme T. Lloyd4, Xiaoli Wang2,3, Yan Wang2,3, Xiaomei Zhang2,3 & Zhonghe Zhou1 Ornithuromorpha is the most inclusive clade containing extant birds but not the Mesozoic Enantiornithes. The early evolutionary history of this avian clade has been advanced with recent discoveries from Cretaceous deposits, indicating that Ornithuromorpha and Enantiornithes are the two major avian groups in Mesozoic. Here we report on a new ornithuromorph bird, Archaeornithura meemannae gen. et sp. nov., from the second oldest avian-bearing deposits (130.7 Ma) in the world. The new taxon is referable to the Hongshanornithidae and constitutes the oldest record of the Ornithuromorpha. However, A. meemannae shows few primitive features relative to younger hongshanornithids and is deeply nested within the Hongshanornithidae, suggesting that this clade is already well established. The new discovery extends the record of Ornithuromorpha by five to six million years, which in turn pushes back the divergence times of early avian lingeages into the Early Cretaceous. 1 Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China. 2 Institue of Geology and Paleontology, Linyi University, Linyi, Shandong 276000, China. 3 Tianyu Natural History Museum of Shandong, Pingyi, Shandong 273300, China. 4 Department of Biological Sciences, Faculty of Science, Macquarie University, Sydney, New South Wales 2019, Australia.