Neuroanatomy of the Visual Pathways
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Distance Learning Program Anatomy of the Human Brain/Sheep Brain Dissection
Distance Learning Program Anatomy of the Human Brain/Sheep Brain Dissection This guide is for middle and high school students participating in AIMS Anatomy of the Human Brain and Sheep Brain Dissections. Programs will be presented by an AIMS Anatomy Specialist. In this activity students will become more familiar with the anatomical structures of the human brain by observing, studying, and examining human specimens. The primary focus is on the anatomy, function, and pathology. Those students participating in Sheep Brain Dissections will have the opportunity to dissect and compare anatomical structures. At the end of this document, you will find anatomical diagrams, vocabulary review, and pre/post tests for your students. The following topics will be covered: 1. The neurons and supporting cells of the nervous system 2. Organization of the nervous system (the central and peripheral nervous systems) 4. Protective coverings of the brain 5. Brain Anatomy, including cerebral hemispheres, cerebellum and brain stem 6. Spinal Cord Anatomy 7. Cranial and spinal nerves Objectives: The student will be able to: 1. Define the selected terms associated with the human brain and spinal cord; 2. Identify the protective structures of the brain; 3. Identify the four lobes of the brain; 4. Explain the correlation between brain surface area, structure and brain function. 5. Discuss common neurological disorders and treatments. 6. Describe the effects of drug and alcohol on the brain. 7. Correctly label a diagram of the human brain National Science Education -

Magnetic Resonance Imaging of Multiple Sclerosis: a Study of Pulse-Technique Efficacy
691 Magnetic Resonance Imaging of Multiple Sclerosis: A Study of Pulse-Technique Efficacy Val M. Runge1 Forty-two patients with the clinical diagnosis of multiple sclerosis were examined by Ann C. Price1 proton magnetic resonance imaging (MRI) at 0.5 T. An extensive protocol was used to Howard S. Kirshner2 facilitate a comparison of the efficacy of different pulse techniques. Results were also Joseph H. Allen 1 compared in 39 cases with high-resolution x-ray computed tomography (CT). MRI revealed characteristic abnormalities in each case, whereas CT was positive in only 15 C. Leon Partain 1 of 33 patients. Milder grades 1 and 2 disease were usually undetected by CT, and in all A. Everette James, Jr.1 cases, the abnormalities noted on MRI were much more extensive than on CT. Cerebral abnormalities were best shown with the T2-weighted spin-echo sequence (TE/TR = 120/1000); brainstem lesions were best defined on the inversion-recovery sequence (TE/TI/TR =30/400/1250). Increasing TE to 120 msec and TR to 2000 msec heightened the contrast between normal and abnormal white matter. However, the signal intensity of cerebrospinal fluid with this pulse technique obscured some abnormalities. The diagnosis of multiple sclerosis continues to be a clinical challenge [1,2). The lack of an objective means of assessment further complicates the evaluation of treatment regimens. Evoked potentials, cerebrospinal fluid (CSF) analysis , and computed tomography (CT) are currently used for diagnosis, but all lack sensitivity and/or specificity. Furthermore, postmortem examinations demonstrate many more lesions than those suggested by clinical means [3). -

Anatomy of the Temporal Lobe
Hindawi Publishing Corporation Epilepsy Research and Treatment Volume 2012, Article ID 176157, 12 pages doi:10.1155/2012/176157 Review Article AnatomyoftheTemporalLobe J. A. Kiernan Department of Anatomy and Cell Biology, The University of Western Ontario, London, ON, Canada N6A 5C1 Correspondence should be addressed to J. A. Kiernan, [email protected] Received 6 October 2011; Accepted 3 December 2011 Academic Editor: Seyed M. Mirsattari Copyright © 2012 J. A. Kiernan. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Only primates have temporal lobes, which are largest in man, accommodating 17% of the cerebral cortex and including areas with auditory, olfactory, vestibular, visual and linguistic functions. The hippocampal formation, on the medial side of the lobe, includes the parahippocampal gyrus, subiculum, hippocampus, dentate gyrus, and associated white matter, notably the fimbria, whose fibres continue into the fornix. The hippocampus is an inrolled gyrus that bulges into the temporal horn of the lateral ventricle. Association fibres connect all parts of the cerebral cortex with the parahippocampal gyrus and subiculum, which in turn project to the dentate gyrus. The largest efferent projection of the subiculum and hippocampus is through the fornix to the hypothalamus. The choroid fissure, alongside the fimbria, separates the temporal lobe from the optic tract, hypothalamus and midbrain. The amygdala comprises several nuclei on the medial aspect of the temporal lobe, mostly anterior the hippocampus and indenting the tip of the temporal horn. The amygdala receives input from the olfactory bulb and from association cortex for other modalities of sensation. -
Corticospinal Fibers
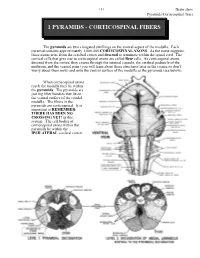
151 Brain stem Pyramids/Corticospinal Tract 1 PYRAMIDS - CORTICOSPINAL FIBERS The pyramids are two elongated swellings on the ventral aspect of the medulla. Each pyramid contains approximately 1,000,000 CORTICOSPINAL AXONS. As the name suggests, these axons arise from the cerebral cortex and descend to terminate within the spinal cord. The cortical cells that give rise to corticospinal axons are called Betz cells. As corticospinal axons descend from the cortex, they course through the internal capsule, the cerebral peduncle of the midbrain, and the ventral pons (you will learn about these structures later in the course so don’t worry about them now) and onto the ventral surface of the medulla as the pyramids (see below). When corticospinal axons reach the medulla they lie within the pyramids. The pyramids are just big fiber bundles that lie on the ventral surface of the caudal medulla. The fibers in the pyramids are corticospinal. It is important to REMEMBER: THERE HAS BEEN NO CROSSING YET! in this system. The cell bodies of corticospinal axons within the pyramids lie within the IPSILATERAL cerebral cortex. Brain stem 152 Pyramids/Corticospinal Tract At the most caudal pole of the pyramids the corticospinal axons cross over the midline and now continue their descent on the contralateral (to the cell of origin) side. This crossover point is called the PYRAMIDAL DECUSSATION. The crossing fibers enter the lateral funiculus of the spinal cord where they are called the LATERAL CORTICOSPINAL TRACT (“corticospinal” is not good enough, you have to call them lateral corticospinal; LCST - remember this one??). LCST axons exit the tract to terminate upon neurons in the spinal cord gray matter along its entire length. -

Brainstem: Structure & Its Mode of Action
Journal of Neurology & Neurophysiology 2021, Vol.12, Issue 3, 521 Opinion Brainstem: Structure & Its Mode of action Karthikeyan Rupani Research Fellow, Tata Medical Centre, India. Corresponding Author* The brainstem is exceptionally little, making up around as it were 2.6 percent of the brain's add up to weight. It has the basic parts of directing cardiac, and Rupani K, respiratory work, making a difference to control heart rate and breathing rate. Research Fellow, Tata Medical Centre, India; It moreover gives the most engine and tactile nerve supply to the confront and E-mail: [email protected] neck by means of the cranial nerves. Ten sets of cranial nerves come from the brainstem. Other parts incorporate the direction of the central apprehensive Copyright: 2021 Rupani K. This is an open-access article distributed under the framework and the body's sleep cycle. It is additionally of prime significance terms of the Creative Commons Attribution License, which permits unrestricted within the movement of engine and tangible pathways from the rest of the use, distribution, and reproduction in any medium, provided the original author brain to the body, and from the body back to the brain. These pathways and source are credited. incorporate the corticospinal tract (engine work), the dorsal column-medial lemniscus pathway and the spinothalamic tract [3]. The primary part of the brainstem we'll consider is the midbrain. The midbrain Received 01 March 2021; Accepted 15 March 2021; Published 22 March 2021 (too known as the mesencephalon) is the foremost prevalent of the three districts of the brainstem. It acts as a conduit between the forebrain over and the pons and cerebellum underneath. -

Quantitative Analysis of Axon Collaterals of Single Pyramidal Cells
Yang et al. BMC Neurosci (2017) 18:25 DOI 10.1186/s12868-017-0342-7 BMC Neuroscience RESEARCH ARTICLE Open Access Quantitative analysis of axon collaterals of single pyramidal cells of the anterior piriform cortex of the guinea pig Junli Yang1,2*, Gerhard Litscher1,3* , Zhongren Sun1*, Qiang Tang1, Kiyoshi Kishi2, Satoko Oda2, Masaaki Takayanagi2, Zemin Sheng1,4, Yang Liu1, Wenhai Guo1, Ting Zhang1, Lu Wang1,3, Ingrid Gaischek3, Daniela Litscher3, Irmgard Th. Lippe5 and Masaru Kuroda2 Abstract Background: The role of the piriform cortex (PC) in olfactory information processing remains largely unknown. The anterior part of the piriform cortex (APC) has been the focus of cortical-level studies of olfactory coding, and asso- ciative processes have attracted considerable attention as an important part in odor discrimination and olfactory information processing. Associational connections of pyramidal cells in the guinea pig APC were studied by direct visualization of axons stained and quantitatively analyzed by intracellular biocytin injection in vivo. Results: The observations illustrated that axon collaterals of the individual cells were widely and spatially distrib- uted within the PC, and sometimes also showed a long associational projection to the olfactory bulb (OB). The data showed that long associational axons were both rostrally and caudally directed throughout the PC, and the intrinsic associational fibers of pyramidal cells in the APC are omnidirectional connections in the PC. Within the PC, associa- tional axons typically followed rather linear trajectories and irregular bouton distributions. Quantitative data of the axon collaterals of two pyramidal cells in the APC showed that the average length of axonal collaterals was 101 mm, out of which 79 mm (78% of total length) were distributed in the PC. -

Visual Cortex in Humans 251
Author's personal copy Visual Cortex in Humans 251 Visual Cortex in Humans B A Wandell, S O Dumoulin, and A A Brewer, using fMRI, and we discuss the main features of the Stanford University, Stanford, CA, USA V1 map. We then summarize the positions and proper- ties of ten additional visual field maps. This represents ã 2009 Elsevier Ltd. All rights reserved. our current understanding of human visual field maps, although this remains an active field of investigation, with more maps likely to be discovered. Finally, we Human visua l cortex comprises 4–6 billion neurons that are organ ized into more than a dozen distinct describe theories about the functional purpose and functional areas. These areas include the gray matter organizing principles of these maps. in the occi pital lobe and extend into the temporal and parietal lobes . The locations of these areas in the The Size and Location of Human Visual intact human cortex can be identified by measuring Cortex visual field maps. The neurons within these areas have a variety of different stimulus response proper- The entirety of human cortex occupies a surface area 2 ties. We descr ibe how to measure these visual field on the order of 1000 cm and ranges between 2 and maps, their locations, and their overall organization. 4 mm in thickness. Each cubic millimeter of cortex contains approximately 50 000 neurons so that neo- We then consider how information about patterns, objects, color s, and motion is analyzed and repre- cortex in the two hemispheres contain on the order of sented in these maps. -

Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex Hans Ten Donkelaar, Nathalie Tzourio-Mazoyer, Jürgen Mai
Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex Hans ten Donkelaar, Nathalie Tzourio-Mazoyer, Jürgen Mai To cite this version: Hans ten Donkelaar, Nathalie Tzourio-Mazoyer, Jürgen Mai. Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex. Frontiers in Neuroanatomy, Frontiers, 2018, 12, pp.93. 10.3389/fnana.2018.00093. hal-01929541 HAL Id: hal-01929541 https://hal.archives-ouvertes.fr/hal-01929541 Submitted on 21 Nov 2018 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. REVIEW published: 19 November 2018 doi: 10.3389/fnana.2018.00093 Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex Hans J. ten Donkelaar 1*†, Nathalie Tzourio-Mazoyer 2† and Jürgen K. Mai 3† 1 Department of Neurology, Donders Center for Medical Neuroscience, Radboud University Medical Center, Nijmegen, Netherlands, 2 IMN Institut des Maladies Neurodégénératives UMR 5293, Université de Bordeaux, Bordeaux, France, 3 Institute for Anatomy, Heinrich Heine University, Düsseldorf, Germany The gyri and sulci of the human brain were defined by pioneers such as Louis-Pierre Gratiolet and Alexander Ecker, and extensified by, among others, Dejerine (1895) and von Economo and Koskinas (1925). -

Quantity Determination and the Distance Effect with Letters, Numbers, and Shapes: a Functional MR Imaging Study of Number Processing
AJNR Am J Neuroradiol 23:193–200, February 2003 Quantity Determination and the Distance Effect with Letters, Numbers, and Shapes: A Functional MR Imaging Study of Number Processing Robert K. Fulbright, Stephanie C. Manson, Pawel Skudlarski, Cheryl M. Lacadie, and John C. Gore BACKGROUND AND PURPOSE: The ability to quantify, or to determine magnitude, is an important part of number processing, and the extent to which language and other cognitive abilities are involved with number processing is an area of interest. We compared activation patterns, reaction times, and accuracy as subjects determined stimulus magnitude by ordering letters, numbers, and shapes. A second goal was to define the brain regions involved in the distance effect (the farther apart numbers are, the faster subjects are at judging which number is larger) and whether this effect depended on stimulus type. METHODS: Functional MR images were acquired in 19 healthy subjects. The order task required the subjects to judge whether three stimuli were in order according to their position in the alphabet (letters), position in the number line (numbers), or size (shapes). In the control (identify task), subjects judged whether one of the three stimuli was a particular letter, number, or shape. Each stimulus type was divided into near trials (quantity difference of three or less) and far trials (quantity difference of at least five) to assess the distance effect. RESULTS: Subjects were less accurate and slower with letters than with numbers and shapes. A distance effect was present with shapes and numbers, as subjects ordered the near trials slower than far trials. No distance effect was detected with letters. -

What's the Connection?
WHAT’S THE CONNECTION? Sharon Winter Lake Washington High School Directions for Teachers 12033 NE 80th Street Kirkland, WA 98033 SYNOPSIS Students elicit and observe reflex responses and distinguish between types STUDENT PRIOR KNOWL- of reflexes. They then design and conduct experiments to learn more about EDGE reflexes and their control by the nervous system. Before participating in this LEVEL activity students should be able to: Exploration, Concept/Term Introduction Phases ■ Describe the parts of a Application Phase neuron and explain their functions. ■ Distinguish between sensory and motor neurons. Getting Ready ■ Describe briefly the See sidebars for additional information regarding preparation of this lab. organization of the nervous system. Directions for Setting Up the Lab General: INTEGRATION Into the Biology Curriculum ■ Make an “X” on the chalkboard for the teacher-led introduction. ■ Health ■ Photocopy the Directions for Students pages. ■ Biology I, II ■ Human Anatomy and Teacher Background Physiology A reflex is an involuntary neural response to a specific sensory stimulus ■ AP Biology that threatens the survival or homeostatic state of an organism. Reflexes Across the Curriculum exist in the most primitive of species, usually with a protective function for ■ Mathematics animals when they encounter external and internal stimuli. A primitive ■ Physics ■ example of this protective reflex is the gill withdrawal reflex of the sea slug Psychology Aplysia. In humans and other vertebrates, protective reflexes have been OBJECTIVES maintained and expanded in number. Examples are the gag reflex that At the end of this activity, occurs when objects touch the sides students will be able to: or the back of the throat, and the carotid sinus reflex that restores blood ■ Identify common reflexes pressure to normal when baroreceptors detect an increase in blood pressure. -

Bilateral Cerebellar Dysfunctions in a Unilateral Meso-Diencephalic Lesion
J Neurol Neurosurg Psychiatry: first published as 10.1136/jnnp.44.4.361 on 1 April 1981. Downloaded from Journal of Neurology, Neurosurgery, and Psychiatry, 1981, 44, 361-363 Short report Bilateral cerebellar dysfunctions in a unilateral meso-diencephalic lesion D VON CRAMON From the Max-Planck-Institute for Psychiatry, Munich, Germany SUMMARY The clinical syndrome of a 65-year-old patient with a slit-shaped right-sided meso- diencephalic lesion was analysed. A cerebellar syndrome with limb-kinetic ataxia, intention tremor and hypotonicity in all extremities as well as ataxic dysarthria was found. The disruption of the two cerebello-(rubro)-thalamic pathways probably explained the signs of bilateral cere- bellar dysfunction. The uncrossed ascending limb of the right, and the crossed one of the left brachium conjunctivum may have been damaged by the unilateral lesion extending between caudal midbrain and dorsal thalamus. Protected by copyright. Most of the fibres which constitute the superior general hospital where neurological examination cerebellar peduncle leave the cerebellum and showed bilateral miosis, convergent strabism, vertical originate in cells of the dentate nucleus but also gaze paresis on upward gaze with gaze-paretic nystag- arise from neurons of the globose and emboli- mus, flaccid sensori-motor hemiparesis with increased stretch reflexes and Babinski sign on the left side, forme nuclei. The crossed ascending fibres of the and dysmetric movements of the right upper extremity. brachia conjunctiva constitute the major outflow The CT scan showed an acute haemorrhage in the from the cerebellum, they form the cerebello- right mesodiencephalic area. On 19 February 1979 (rubro)-thalamic and dentato-thalamic tracts.' the patient was admitted to our department. -

A Model of Accommodative-Pupillary Dynamics Stanley Gordon Day Iowa State University
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1969 A model of accommodative-pupillary dynamics Stanley Gordon Day Iowa State University Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Electrical and Electronics Commons Recommended Citation Day, Stanley Gordon, "A model of accommodative-pupillary dynamics " (1969). Retrospective Theses and Dissertations. 4649. https://lib.dr.iastate.edu/rtd/4649 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. This dissertation has been microâhned exactly as received 69-15,607 DAY, Stanley Gordon, 1939- A MODEL OF ACCOMMODATIVE-PUPILLARY DYNAMICS. Iowa State University, Ph.D., 1969 Engineering, electrical University Microfilms, Inc., Ann Arbor, Michigan ®Copyright by STANLEY GORDON DAY 1969 A MODEL OF ACCOMMODATIVE-PUPILLARY DYNAMICS by Stanley Gordon Day A Dissertation Submitted to the Graduate Faculty in Partial Fulfillment of The Requirements for the Degree of DOCTOR OF PHILOSOPHY Major Subject : Electrical Engineering Approved: Signature was redacted for privacy. In Charge of Major Work Signature was redacted for privacy. Head of Major Department Signature was redacted for privacy. Dea^ of Gradulate College Iowa State University Of Science and Technology Ames, Iowa 1969 il TABLE OF CONTENTS Page DEDICATION iii INTRODUCTION 1 REVIEW OF LITERATURE 4 EQUIPMENT AND METHODS 22 RESULTS AND DISCUSSION 47 SUÎ4MARY AND CONCLUSIONS 60 BIBLIOGRAPHY 62 ACKNOWLEDGEMENTS 68 APPENDIX 69 i iii DEDICATION This dissertation is dedicated to Sandra R.