Adaptive Radiation Into Ecological Niches with Eruptive Dynamics: A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

ARTHROPOD COMMUNITIES and PASSERINE DIET: EFFECTS of SHRUB EXPANSION in WESTERN ALASKA by Molly Tankersley Mcdermott, B.A./B.S
Arthropod communities and passerine diet: effects of shrub expansion in Western Alaska Item Type Thesis Authors McDermott, Molly Tankersley Download date 26/09/2021 06:13:39 Link to Item http://hdl.handle.net/11122/7893 ARTHROPOD COMMUNITIES AND PASSERINE DIET: EFFECTS OF SHRUB EXPANSION IN WESTERN ALASKA By Molly Tankersley McDermott, B.A./B.S. A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of Master of Science in Biological Sciences University of Alaska Fairbanks August 2017 APPROVED: Pat Doak, Committee Chair Greg Breed, Committee Member Colleen Handel, Committee Member Christa Mulder, Committee Member Kris Hundertmark, Chair Department o f Biology and Wildlife Paul Layer, Dean College o f Natural Science and Mathematics Michael Castellini, Dean of the Graduate School ABSTRACT Across the Arctic, taller woody shrubs, particularly willow (Salix spp.), birch (Betula spp.), and alder (Alnus spp.), have been expanding rapidly onto tundra. Changes in vegetation structure can alter the physical habitat structure, thermal environment, and food available to arthropods, which play an important role in the structure and functioning of Arctic ecosystems. Not only do they provide key ecosystem services such as pollination and nutrient cycling, they are an essential food source for migratory birds. In this study I examined the relationships between the abundance, diversity, and community composition of arthropods and the height and cover of several shrub species across a tundra-shrub gradient in northwestern Alaska. To characterize nestling diet of common passerines that occupy this gradient, I used next-generation sequencing of fecal matter. Willow cover was strongly and consistently associated with abundance and biomass of arthropods and significant shifts in arthropod community composition and diversity. -

Forestry Department Food and Agriculture Organization of the United Nations
Forestry Department Food and Agriculture Organization of the United Nations Forest Health & Biosecurity Working Papers OVERVIEW OF FOREST PESTS ROMANIA January 2007 Forest Resources Development Service Working Paper FBS/28E Forest Management Division FAO, Rome, Italy Forestry Department DISCLAIMER The aim of this document is to give an overview of the forest pest1 situation in Romania. It is not intended to be a comprehensive review. The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. © FAO 2007 1 Pest: Any species, strain or biotype of plant, animal or pathogenic agent injurious to plants or plant products (FAO, 2004). Overview of forest pests - Romania TABLE OF CONTENTS Introduction..................................................................................................................... 1 Forest pests and diseases................................................................................................. 1 Naturally regenerating forests..................................................................................... 1 Insects ..................................................................................................................... 1 Diseases................................................................................................................ -

Forest Insect and Disease Conditions in the United States 2000
United States Department Forest Insect and Of Agriculture Forest Service Disease Conditions Forest Health Protection in the United States March 2002 2000 Healthy Forests Make A World of Difference United States Department Of Agriculture Forest Insect and Forest Service Disease Conditions Forest Health Protection in the United States March 2002 2000 PREFACE This is the 50th annual report prepared by the U.S. • seed orchard insects and diseases; Department of Agriculture Forest Service (USDA • nursery insects and diseases; and Forest Service) of the insect and disease conditions of • abiotic damage. the Nation's forests. This report responds to direction in the Cooperative Forestry Assistance Act of 1978, as These categories are listed in the table of contents; amended, to conduct surveys and report annually on there is no index. insect and disease conditions of major national significance. Insect and disease conditions of local The information in this report is provided by the Forest importance are reported in regional and State reports. Health Protection Program of the USDA Forest Service. This program serves all Federal lands, The report describes the extent and nature of insect- including the National Forest System and the lands and disease-caused damage of national significance in administered by the Departments of Defense and 2000. As in the past, selected insect and disease Interior. Service is also provided to tribal lands. The conditions are highlighted in the front section of the program provides assistance to private landowners report. Maps are provided for some pests showing through the State foresters. A key part of the program affected counties in the East and affected areas in the is detecting and reporting insect and disease epidemics West. -

Scaffolds and Crop Development R U I T U N a July 31, 2000 VOLUME 9, No
Update on Pest Management scaffolds and Crop Development R U I T u N A July 31, 2000 VOLUME 9, No. 20 Geneva, NY PLEASE DON'T They are also very expensive. EAT THESE DAISIES This regular annual article used (Dave Kain & to state that the four most common Art Agnello, botanicals available for use in fruit Entomology, crops today were rotenone, pyre- Geneva) thrin, sabadilla and ryania. Unfortu nately, for those who found them use ❖ ❖ Naturally occuring pesticides ful, sabadilla and ryania are no longer on that are derived from plants or plant parts are the list due to voluntary cancellation of their commonly referred to as “botanicals”. Botani- registrations. To round out the article, we’ll sub cals have been around for quite a while. Along stitute information on a few, newer, natural ma with arsenicals and other inorganic pesticides, terials that, while not technically botanicals, kind they were pretty commonly used before the of fit the category. Information on these products advent of the synthetic, organic pesticides ren appears in the 2000 Tree-Fruit Recommenda dered them “obsolete”. From time to time they’re tions (pp. 24-26). re-examined for various reasons and may be familiar. Botanicals are of interest to those ROTENONE Rotenone is derived from the root concerned with pest management for a variety of of various plants of the Derris or Lonchocarpus reasons. They are generally less toxic to the species from Southeast Asia, Central and South applicator than many synthetic pesticides. They America. It is available as at least 118 formu may be acceptable in the organic market where lated products from a large number of manufac synthetic pesticides are not. -

Status and Protection of Globally Threatened Species in the Caucasus
STATUS AND PROTECTION OF GLOBALLY THREATENED SPECIES IN THE CAUCASUS CEPF Biodiversity Investments in the Caucasus Hotspot 2004-2009 Edited by Nugzar Zazanashvili and David Mallon Tbilisi 2009 The contents of this book do not necessarily reflect the views or policies of CEPF, WWF, or their sponsoring organizations. Neither the CEPF, WWF nor any other entities thereof, assumes any legal liability or responsibility for the accuracy, completeness, or usefulness of any information, product or process disclosed in this book. Citation: Zazanashvili, N. and Mallon, D. (Editors) 2009. Status and Protection of Globally Threatened Species in the Caucasus. Tbilisi: CEPF, WWF. Contour Ltd., 232 pp. ISBN 978-9941-0-2203-6 Design and printing Contour Ltd. 8, Kargareteli st., 0164 Tbilisi, Georgia December 2009 The Critical Ecosystem Partnership Fund (CEPF) is a joint initiative of l’Agence Française de Développement, Conservation International, the Global Environment Facility, the Government of Japan, the MacArthur Foundation and the World Bank. This book shows the effort of the Caucasus NGOs, experts, scientific institutions and governmental agencies for conserving globally threatened species in the Caucasus: CEPF investments in the region made it possible for the first time to carry out simultaneous assessments of species’ populations at national and regional scales, setting up strategies and developing action plans for their survival, as well as implementation of some urgent conservation measures. Contents Foreword 7 Acknowledgments 8 Introduction CEPF Investment in the Caucasus Hotspot A. W. Tordoff, N. Zazanashvili, M. Bitsadze, K. Manvelyan, E. Askerov, V. Krever, S. Kalem, B. Avcioglu, S. Galstyan and R. Mnatsekanov 9 The Caucasus Hotspot N. -

Chlorantraniliprole: Reduced-Risk Insecticide for Controlling Insect Pests of Woody Ornamentals with Low Hazard to Bees
242 Redmond and Potter: Acelepryn Control of Horticultural Pests Arboriculture & Urban Forestry 2017. 43(6):242–256 Chlorantraniliprole: Reduced-risk Insecticide for Controlling Insect Pests of Woody Ornamentals with Low Hazard to Bees Carl T. Redmond and Daniel A. Potter Abstract. Landscape professionals need target-selective insecticides for managing insect pests on flowering woody orna- mentals that may be visited by bees and other insect pollinators. Chlorantraniliprole, the first anthranilic diamide insec- ticide registered for use in urban landscapes, selectively targets the receptors that regulate the flow of calcium to control muscle contraction in caterpillars, plant-feeding beetles, and certain other phytophagous insects. Designated a reduced- risk pesticide by the United States Environmental Protection Agency, it has a favorable toxicological and environmental profile, including very low toxicity to bees and most types of predatory and parasitic insects that contribute to pest sup- pression. Chlorantraniliprole has become a mainstay for managing turfgrass pests, but little has been published concern- ing its performance against the pests of woody ornamentals. Researchers evaluated it against pests spanning five different orders: adult Japanese beetles, evergreen bagworm, eastern tent caterpillar, bristly roseslug sawfly, hawthorn lace bug, ole- ander aphid, boxwood psyllid, oak lecanium scale (crawlers), and boxwood leafminer, using real-world exposure scenarios. Chlorantraniliprole’s efficacy, speed of control, and residual activity as a foliar spray for the leaf-chewing pests was as good, or better, than provided by industry standards, but sprays were ineffective against the sucking pests (lace bugs, aphids, or scales). Basal soil drenches in autumn or spring failed to systemically control boxwood psyllids or leafminers, but autumn drenches did suppress roseslug damage and Japanese beetle feeding the following year. -

Revista Chilena De Entomología
Rev. Chilena Ent. 1990, 18: 5-7 AMETASTEGIA GLABRATA (FALLEN) ESPECIE FITÓFAGA INTRODUCIDA A CHILE (HYMENOPTERA: TENTHREDINIDAE) Roberto Carrillo Ll.\ Nelly Mundaca B.' y Ernesto Cisternas A.^ RESUMEN Se señala la presencia en Chile del tentredínido Ametastegia glabrata (Fallen), especie holártica de hábitos polífagos, que se alimenta de diversas plantas cultivadas y malezas. Este insecto ha sido encontrado en diversas localidades de la X Región. Se entregan algunos antecedentes de su biología y de sus plantas hospederas. ABSTRACT The presence in Chile of the holoartic sawfly Ametastegia glabrata (Fallen) is reported. It is a polypha- gous insect, that feed on different cultivated plants and weeds. The insect has been reported in different localities of the X'^ Región. Aspects of its biology and hosts are reported. INTRODUCCIÓN de crianza (± 19°C). En el mes de junio se obtuvo un adulto en Remehue, y a partir de En el otoño de 1987, en la comuna de La fines de octubre, en la Universidad Austral de Unión, en el lugar denominado Choroico, se Chile. Los adultos fueron enviados al Dr. Da- observó en una plantación de frambuesa de vid Smith, del U.S. National Museum, quien primer año, cv. Heritage, algunas cañas caídas identificó la especie como A. glabrata Fallen. sobre el suelo y otras se encontraban dobladas en forma de V invertida. Examinadas estas Clasifícación cañas se observó, en su interior o en el lugar en el cual se había producido el quiebre, una La especie A. glabrata pertenece a la familia larva de tendredínido de color verde. Revisio- Tenthredinidae, subfamilia Allantinae y tribu nes efectuadas en la misma temporada en Empriini. -

Ana Kurbalija PREGLED ENTOMOFAUNE MOČVARNIH
SVEUČILIŠTE JOSIPA JURJA STROSSMAYERA U OSIJEKU I INSTITUT RUĐER BOŠKOVI Ć, ZAGREB Poslijediplomski sveučilišni interdisciplinarni specijalisti čki studij ZAŠTITA PRIRODE I OKOLIŠA Ana Kurbalija PREGLED ENTOMOFAUNE MOČVARNIH STANIŠTA OD MEĐUNARODNOG ZNAČENJA U REPUBLICI HRVATSKOJ Specijalistički rad Osijek, 2012. TEMELJNA DOKUMENTACIJSKA KARTICA Sveučilište Josipa Jurja Strossmayera u Osijeku Specijalistički rad Institit Ruđer Boškovi ć, Zagreb Poslijediplomski sveučilišni interdisciplinarni specijalisti čki studij zaštita prirode i okoliša Znanstveno područje: Prirodne znanosti Znanstveno polje: Biologija PREGLED ENTOMOFAUNE MOČVARNIH STANIŠTA OD ME ĐUNARODNOG ZNAČENJA U REPUBLICI HRVATSKOJ Ana Kurbalija Rad je izrađen na Odjelu za biologiju, Sveučilišta Josipa Jurja Strossmayera u Osijeku Mentor: izv.prof. dr. sc. Stjepan Krčmar U ovom radu je istražen kvalitativni sastav entomof aune na četiri močvarna staništa od me đunarodnog značenja u Republici Hrvatskoj. To su Park prirode Kopački rit, Park prirode Lonjsko polje, Delta rijeke Neretve i Crna Mlaka. Glavni cilj specijalističkog rada je objediniti sve objavljene i neobjavljene podatke o nalazima vrsta kukaca na ova četiri močvarna staništa te kvalitativno usporediti entomofau nu pomoću Sörensonovog indexa faunističke sličnosti. Na području Parka prirode Kopački rit utvrđeno je ukupno 866 vrsta kukaca razvrstanih u 84 porodice i 513 rodova. Na području Parka prirode Lonjsko polje utvrđeno je 513 vrsta kukaca razvrstanih u 24 porodice i 89 rodova. Na području delte rijeke Neretve utvrđeno je ukupno 348 vrsta kukaca razvrstanih u 89 porodica i 227 rodova. Za područje Crne Mlake nije bilo dostupne literature o nalazima kukaca. Velika vrijednost Sörensonovog indexa od 80,85% ukazuje na veliku faunističku sličnost između faune obada Kopačkoga rita i Lonjskoga polja. Najmanja sličnost u fauni obada utvrđena je između močvarnih staništa Lonjskog polja i delte rijeke Neretve, a iznosi 41,37%. -
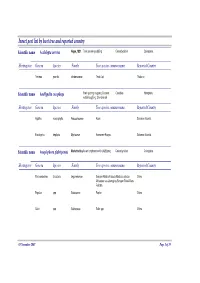
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae -

Bees, Wasps & Ants
Sheringham and Beeston Regis Commons SSSI / SAC FAUNA: Hymenoptera INSECTA (Pterygota) Family/Order English Name. Scientific Name. Authority. Grid Ref. Tetrad/ Last Km sq. Common. Record. HYMENOPTERA. PAMPHILIDAE: Sawfly. Pamphilius inanitus (Villers, 1789) TG1642 1987? (Bees, Wasps and Ants) ARGIDAE: Elm Zig-zag Sawfly. Aproceros leucopoda Takeuchi, 1939 TG1642 14R/B 2020 Bramble Sawfly. Arge cyaneocrocea (Forster, 1771) TG1642 2016 Sawfly. Arge gracilicornis (Klug, 1814 ) TG1642 1987? CIMBICIDAE: Honeysuckle Sawfly. Abia lonicerae (Linnaeus) TG1641 14Q/B 2015 Club-horned Sawfly. Abia sericera (Linnaeus) TG1642 14R/B 2014 Club-horned Sawfly. Zaraea fasciata Linnaeus, 1758 TG1641/42 14R,14Q/B 2014 Birch Sawfly. Cimbex femoratus (Linnaeus, 1758) TG1642 14R/B 2017 SIRICIDAE: Greater Horntail Wasp. Urocerus gigas (Linnaeus, 1758) TG1642 14R/S 1992 CEPHIDAE: Sawfly. Calameuta pallipes (Klug, 1803) TG1642 1987? TENTHREDINIDAE: Willow Sawfly. Pontania proxima (Lepeletier, 1823) TG1642 14R/BS 2009 Willow Sawfly. Eupontania pedunculi (Hartig, 1837) TG1642 14R/B 1999 Willow Sawfly. Eupontainia viminalis (Linnaeus, 1758) TG1642 14R/B 2002 Willow Sawfly. Pontainia bridgemanii (Cameron, 1883) TG1642 14R/B 1999 Sawfly. Caliroa annulipes (Klug, 1816) TG1642 14R/S 2002 Hazel Sawfly. Craesus septentrionalis (Linnaeus, 1758) TG1641 14Q/B 2017 Sawfly. Blennocampa phyllocolpa Viitasaari & Vikberg, 1985 TG1642/41 14R,14Q/B 2003 Sawfly. Selandria serva (Fabricius, 1793) TG1642 14R/B 2013 Sawfly. Aneugmenus padi (Linnaeus, 1761) TG1642 1987? Bracken Sawfly. Strongylogaster multifasciata (Geoffroy, 1785) TG1642 14R/BS 2020 Sawfly. Dichrodolerus vestigialis (Klug, 1818) TG1642 1996 Sawfly. Dolerus germanicus (Fabricius, 1775) TG1642 1987? Sawfly. Eutomostethus ephippium (Panzer, 1798) TG1642 14R/BS 2020 Sawfly. Poodolerus aeneus Hartig, 1837 TG1642 1987? Sawfly. Dolerus brevitarus Hartig TG1642 1987? Sawfly. -

SPECIES of PHYTOPHAGOUS INSECTS ASSOCIATED with STRAWBERRIES in LATVIA Valentîna Petrova, Lîga Jankevica, and Ineta Samsone
PROCEEDINGS OF THE LATVIAN ACADEMY OF SCIENCES. Section B, Vol. 67 (2013), No. 2 (683), pp. 124–129. DOI: 10.2478/prolas-2013-0019 SPECIES OF PHYTOPHAGOUS INSECTS ASSOCIATED WITH STRAWBERRIES IN LATVIA Valentîna Petrova, Lîga Jankevica, and Ineta Samsone Institute of Biology, University of Latvia, Miera ielâ 3, Salaspils, LV-2169, LATVIA [email protected] Communicated by Viesturs Melecis The aim of the present study was to describe the phytophagous insect fauna of strawberries in Latvia. This study was carried out in 2000–2004 on strawberry plantations in Tukums, Rîga, Do- bele, and Limbaþi districts. Insects were collected from strawberry fields by pitfall trapping, sweep netting and leaf sampling methods. A total of 137 insect species belonging to seven orders and 41 families were identified to species. Of the phytophagous insects, the order Orthoptera was represented by one species, other orders by a larger number of species: Hymenoptera (3), Dip- tera (16), Lepidoptera (20), Thysanoptera (21), Hemiptera (39), and Coleoptera (37). Of the re- corded insects, 48 species have a status of general strawberry pests. Key words: Fragaria × ananassa, strawberry pests, insect diversity. INTRODUCTION planted in rows with 30-cm distance between plants and 100-cm distance between rows. In the period from June 1 to Strawberries are one of the commercially important crop September 30 in 2000–2004, random sweep netting plants in Latvia. Harmful phytophagous insect species on (monthly) and leaf sampling (twice a month) were performed strawberry have been studied during the period between for general collection of homopterans, thysanopterans, 1928 and 1989. Thirty-four phytophagous insect species lepidopterans and hemipterans. -

Technical Series, No
' ' Technical Series, No. 20, Part II. U. S. DEPARTMENT OF AGRICULTURE, BXJRE^TJ OK' TClSrTOM:OIL.OG^Y. L, 0. HOWARD, Entomologist and Chief of Bureau. TECHNICAL PAPERS ON MISCELLANEOUS .FOREST INSECTS. II. THE GENOTYPES OF THE SAWFLIES AND WOODWASPS, OR THE SUPERFAMILY TENTHKEDINOIDEA. S. A. ROHWER, Agent and Expert. Issued M.\rch 4, 1911. WASHINGTON: GOVERNMENT PRINTING OFFICE. 1911. Technical Series, No. 20, Part II. U. S. DEPARTMENT OF AGRICULTURE. L. 0. HOWARD, Entomologist and Chief of Bureau. TECHNICAL PAPERS ON MISCELLANEOUS FOREST INSECTS. II. THE GENOTYPES OF THE SAWFLIES AND WOODWASPS, OR THE SUPERFAMILY TENTHREDINOIDEA. BY S. A. ROHWER, Agent and Expert. Issued Makch 4, 1911. WASHINGTON: GOVERNMENT PRINTING OFFICE. 1911. B UREA U OF ENTOMOLOGY. L. O. Howard, Entomologist and Chief of Bureau. C. L. Marlatt, Entomologist and Acting Chief in Absence of Chief. R. S. Clifton, Executive Assistant. W. F. Tastet, Chief Clerk. F. H. Chittenden, in charge of truck crop and stored product insect investigations. A. D. Hopkins, in charge offorest insect investigations. W. D. Hunter, in charge of southern field crop insect investigations. F. M. Webster, in charge of cereal and forage insect investigations. A. L. Quaintance, in charge of deciduous fruit insect investigations. E. F. Phillips, in charge of bee culture. D. M. Rogers, in charge of preventing spread of moths, field -work. RoLLA P. Currie, in charge of editorial work. Mabel Colcord, librarian. , Forest Insect Investigations. A. D. Hopkins, in charge. H. E. Burke, J. L. Webb, Josef Brunner, S. A. Rohwer, T. E. Snyder, W. D. Edmonston, W. B. Turner, agents and experts.