Hemichordata and Chordata Are Two Deuterostome Phyla
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ascidian Cannibalism Correlates with Larval Behavior and Adult Distribution
FAU Institutional Repository http://purl.fcla.edu/fau/fauir This paper was submitted by the faculty of FAU’s Harbor Branch Oceanographic Institute. Notice: ©1988 Elsevier Ltd. The final published version of this manuscript is available at http://www.sciencedirect.com/science/journal/00220981 and may be cited as: Young, C. M. (1988). Ascidian cannibalism correlates with larval behavior and adult distribution. Journal of Experimental Marine Biology and Ecology, 117(1), 9-26. doi:10.1016/0022-0981(88)90068-8 J. Exp. Mar. Bioi. £Col., 1988, Vol. 117, pp. 9-26 9 Elsevier JEM 01042 Ascidian cannibalism correlates with larval behavior and adult distribution Craig M. Young Department ofLarval Ecology. Harbor Branch Oceanographic Institution, Fort Pierce, Florida. U.S.A. (Received 24 March 1987; revision received 9 December 1987; accepted 22 December 1987) Abstract: In the San Juan Islands, Washington, solitary ascidians .that occur in dense monospecific aggregations demonstrate gregarious settlement as larvae, whereas species that occur as isolated individuals do not. All gregarious species reject their own eggs and larvae as food, but nongregarious species consume conspecific eggs and larvae. Moreover, the rejection mechanism is species-specific in some cases. Correla tion analysis suggests that species specificity of the rejection response has a basis in siphon diameter, egg density, and larval size, but not in number of oral tentacles, or tentacle branching. One strongly cannibalistic species, Corella inflata Huntsman, avoids consuming its own eggs and newly released tadpoles by a unique brooding mechanism that involves floating eggs, negative geotaxis after hatching, and adult orientation. Key words: Ascidian; Cannibalism; Distribution; Larva; Settlement behavior INTRODUCTION Many sessile marine invertebrates, including filter-feeders such as mussels, oysters, barnacles and ascidians, occur in discrete, dense aggregations. -
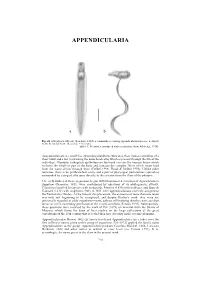
Appendicularia of CTAW
APPENDICULARIA APPENDICULARIA a b Fig. 26. Oikopleura albicans (Leuckart, 1854), a commonly occurring appendicularian species: a, dorsal view; b, lateral view. (Scale bar = 0.1 mm). [after T. Prentiss, reproduced with permission, from Alldredge 1976] Appendicularians are small free swimming planktonic tunicates, their bodies consisting of a short trunk and a tail (containing the notochord cells) which is present through the life of the individual. Glandular (oikoplast) epithelium on the trunk secretes the mucous house which encloses the whole or part of the body and contains the complex filters which strain food from the water driven through them (Deibel 1998; Flood & Deibel 1998). Unlike other tunicates, there is no peribranchial cavity and a pair of pharyngeal perforations (spiracles) surrounded by a ring of cilia open directly to the exterior from the floor of the pharynx. The early studies of these organisms, begun with Chamisso's description of Appendicularia flagellum Chamisso, 1821, were confounded by questions of its phylogenetic affinity. Chamisso classified his species with medusoids, Mertens (1830) with molluscs, and Quoy & Gaimard (1833) with zoophytes. Only in 1851 were appendicularians correctly assigned to the Tunicata by Huxley. At the time of this placement, the existence of more than one taxon was only just beginning to be recognised, and despite Huxley's work, they were not universally regarded as adult organisms—some authors still insisting that they were ascidian larvae or a free swimming generation of the sessile ascidians (Fenaux 1993). Subsequently, these questions were resolved by the work of Fol (1872) on material from the Straits of Messina, which forms the basis of later studies on the large collections of the great expeditions of the 19th century that revealed their true diversity in the oceanic plankton. -

Eurochordata and Cephalochordata of Persian Gulf & Oman See
Subphylum Urochordata : Tunicates Class Ascidacea Class Larvacea Class Thaliacea Subphylum Cephalochordata : Amphioxus Phylum Chordata : 1 - Subphylum Urochordata 2 Subphylum Cephalochordata (Subphylum Urochordata : Tunicates ) Class Ascidiacea Class Larvacea Class Thaliacea (Subphylum Cephalochordata ) Class Ascidiacea Ascidia Kowalevsky Patricia Kott D. lane David George & Gordon Paterson Herdmania momus savigny Herdmania momus kiamanensis Didemnum candidum savigny Phallusia nigra savigny Styela canopus savigny Class Ascidiacea : Order : Aplousobranchia Family : Polyclinidae : Aplidium cf. rubripunctum C.Monniot & F. Monniot 1997 Polyclinum constellatum Savigny, 1816 Family Didemnidae Didemnum candidum Savigny, 1816 Didemnum cf. granulatum Tokioka, 1954 Didemnum obscurum F. Monniot 1969 Didemnum perlucidum F. Monniot, 1983 Didemnum sp. Didemnum yolky C.Monniot & F. Monniot 1997 Diplosoma listerianum Milne Edwards, 1841 Order : Phlebobranchia Family : Ascidiidae Ascidia sp. Phallusia julinea Sluiter, 1915 Phallusia nigra Savigny, 1816 Order : Stolidobranchia Family : Styelidae Botryllus gregalis Sluiter, 1898 Botryllus niger Herdman, 1886 Botryllus sp. Eusynstyela cf. hartmeyeri Michaelsen, 1904 Polyandrocarpa sp. Styela canopus Savigny, 1816 Symplegma bahraini C.Monniot & F. Monniot 1997 Symplegma brakenhielmi Michaelsen, 1904 Family pyuridae Herdmania momus Savigny, 1816 Pyura curvigona Tokioka, 1950 Pyurid sp. Class Larvacea Larva Seymour Sewell Phylum Chordata Subphylum Urochordata * Class Larvacea Order Copelata Family Fritillariidae -

The Origins of Chordate Larvae Donald I Williamson* Marine Biology, University of Liverpool, Liverpool L69 7ZB, United Kingdom
lopmen ve ta e l B Williamson, Cell Dev Biol 2012, 1:1 D io & l l o l g DOI: 10.4172/2168-9296.1000101 e y C Cell & Developmental Biology ISSN: 2168-9296 Research Article Open Access The Origins of Chordate Larvae Donald I Williamson* Marine Biology, University of Liverpool, Liverpool L69 7ZB, United Kingdom Abstract The larval transfer hypothesis states that larvae originated as adults in other taxa and their genomes were transferred by hybridization. It contests the view that larvae and corresponding adults evolved from common ancestors. The present paper reviews the life histories of chordates, and it interprets them in terms of the larval transfer hypothesis. It is the first paper to apply the hypothesis to craniates. I claim that the larvae of tunicates were acquired from adult larvaceans, the larvae of lampreys from adult cephalochordates, the larvae of lungfishes from adult craniate tadpoles, and the larvae of ray-finned fishes from other ray-finned fishes in different families. The occurrence of larvae in some fishes and their absence in others is correlated with reproductive behavior. Adult amphibians evolved from adult fishes, but larval amphibians did not evolve from either adult or larval fishes. I submit that [1] early amphibians had no larvae and that several families of urodeles and one subfamily of anurans have retained direct development, [2] the tadpole larvae of anurans and urodeles were acquired separately from different Mesozoic adult tadpoles, and [3] the post-tadpole larvae of salamanders were acquired from adults of other urodeles. Reptiles, birds and mammals probably evolved from amphibians that never acquired larvae. -

The Ascidian Styela Plicata As a Potential Bioremediator of Bacterial and Algal Contamination of Marine Estuarine Waters
THE ASCIDIAN STYELA PLICATA AS A POTENTIAL BIOREMEDIATOR OF BACTERIAL AND ALGAL CONTAMINATION OF MARINE ESTUARINE WATERS by Lisa Denham Draughon A Dissertation Submitted to the Faculty of The Charles E. Schmidt College of Science In Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy Florida Atlantic University Boca Raton, Florida May, 2010 Copyright by Lisa Denham Draughon 2010 ii ACKNOWLEDGEMENTS I wish to express my sincerest gratitude to those who made this research possible; Ocean Restoration Corporation Associates (ORCA), the Link Foundation, and Florida Atlantic University (FAU) for their financial fellowships, and for the many hands that played a part in getting it all done. A special thank you goes to my committee members; Dr. James X. Hartmann (chairperson) and Dr. David Binninger of FAU, Dr. Ed Proffitt and Dr. John Scarpa of Harbor Branch Oceanographic Institute at FAU, and Dr. Ray Waldner of Palm Beach Atlantic University. You were all such an integral part of helping me achieve this goal. Special thanks also go to my friend Karen Halloway- Adkins who was always willing to help locate specimens and provide equipment when she had it. Mike Calinski of ORCA made trips from the Gulf Coast to deliver specimens to me. Sherry Reed of the Smithsonian Marine Station in Ft. Pierce was extremely helpful by keeping an eye out for the tunicates and at times gathering them for me. Dr. Patricia Keating was invaluable for her help in flow-cytometry analysis. Dr. Peter McCarthy of Harbor Branch Oceanographic Institute at FAU provided the bacteria for the filtration rate experiment. -

Towards an Understanding of Salp Swarm Dynamics. ICES CM 2002/N
CM 2002/ N:12 Towards an understanding of salp swarm dynamics by Patricia Kremer ABSTRACT Species of salps are characterized by intermittent blooms. Several studies have documented the importance of physical processes both in providing seed stocks of salps and creating an environment that is favorable for the rapid increase of salp populations. Although salps are typically oceanic, most observations of bloom dynamics have been made in more accessible inshore waters, so it is difficult to assess how frequent and widespread these swarms are. A review and comparison of existing data is helping to define geographic “hot spots” for salp blooms as well as the necessary physical and biological precursors. Although at this point, the review is far from complete, several generalities are emerging. The details of the physical forcing functions vary, but the overall physical regime seems to require a region of pulsed mixing of oceanic water that results in a relatively high standing stock of autotrophs. For a salp bloom to occur, there also needs to be an adequate seed population of salps and sufficient sustained primary production to support the biomass of the salp population as the bloom develops. As non-selective filter feeders, salps are able to remove a wide range of particulates from the water column, transforming the undigested portion into fast sinking feces. Therefore, when salps occur at high densities, the water is characterized by low abundance of other plankton, with obvious trophic implications. Patricia Kremer: Marine Sciences, University of Connecticut, Groton CT 06340-6097, USA [tel: +1 860 405 9140; fax: +1 860 405 9153; e-mail [email protected]] INTRODUCTION Salps are holoplanktonic grazers that have a life history, feeding biology, and population dynamics that contrasts sharply to copepods and other crustacean zooplankton (Madin and Deibel 1998). -

Field and Laboratory Studies on Four Species of Sea Squirts and Their
Global Journal of Science Frontier Research: B Chemistry Volume 16 Issue 2 Version 1.0 Year 2016 Type : Double Blind Peer Reviewed International Research Journal Publisher: Global Journals Inc. (USA) Online ISSN: 2249-4626 & Print ISSN: 0975-5896 Field and Laboratory Studies on Four Species of Sea Squirts and their Larvae By Gaber Ahmed Saad & Abdullah Bedeer Hussein Dammam University, Saudi Arabia Abstract- The aim of this study was to characterize adult distribution with respect to light and analyze ovary contents in the four seasons of the year. The swimming behavior of Ciona intestinalis, Molgula manhattensis, Ascidella aspersa and Phallusia mammilata larvae against certain abiotic factors were commented. For the field data on adult distributions, one-way analysis of variance (ANOVA) was applied to test for differences in adult orientation, with surface orientation as a fixed factor. Two adult species (Ciona intestinalis and Molgula manhattensis) showed no orientation with respect to light while in the other two species (Ascidella aspersa and Phallusia mammilata) light exerted a significant effects on the orientation and density of individuals. To evaluate among the different species the level of gregariousness found in the field, the number of individuals per clump for each species has been compared using one-way ANOVA, with species as a fixed factor. Artificial heterologous inseminations were carried out. Keywords: field data - gregariousness - heterologous inseminations -adult orientation - larval settlement – phototaxis - geotaxis. GJSFR-B Classification : FOR Code: 069999 FieldandLaboratoryStudiesonFourSpeciesofSeaSquirtsandtheirLarvae Strictly as per the compliance and regulations of : © 2016. Gaber Ahmed Saad & Abdullah Bedeer Hussein. This is a research/review paper, distributed under the terms of the Creative Commons Attribution-Noncommercial 3.0 Unported License http://creativecommons.org/licenses/by-nc/3.0/), permitting all non commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. -

OREGON ESTUARINE INVERTEBRATES an Illustrated Guide to the Common and Important Invertebrate Animals
OREGON ESTUARINE INVERTEBRATES An Illustrated Guide to the Common and Important Invertebrate Animals By Paul Rudy, Jr. Lynn Hay Rudy Oregon Institute of Marine Biology University of Oregon Charleston, Oregon 97420 Contract No. 79-111 Project Officer Jay F. Watson U.S. Fish and Wildlife Service 500 N.E. Multnomah Street Portland, Oregon 97232 Performed for National Coastal Ecosystems Team Office of Biological Services Fish and Wildlife Service U.S. Department of Interior Washington, D.C. 20240 Table of Contents Introduction CNIDARIA Hydrozoa Aequorea aequorea ................................................................ 6 Obelia longissima .................................................................. 8 Polyorchis penicillatus 10 Tubularia crocea ................................................................. 12 Anthozoa Anthopleura artemisia ................................. 14 Anthopleura elegantissima .................................................. 16 Haliplanella luciae .................................................................. 18 Nematostella vectensis ......................................................... 20 Metridium senile .................................................................... 22 NEMERTEA Amphiporus imparispinosus ................................................ 24 Carinoma mutabilis ................................................................ 26 Cerebratulus californiensis .................................................. 28 Lineus ruber ......................................................................... -

Histone H1 and the Origin of Protamines
Histone H1 and the origin of protamines John D. Lewis*†,Nu´ ria Saperas†‡, Yue Song*, Maria Jose Zamora§, Manel Chiva§, and Juan Ausio´ *¶ *Department of Biochemistry and Microbiology, University of Victoria, P.O. Box 3055, Petch Building, Victoria, BC, Canada V8W 3P6; ‡Departament d’Enginyeria Quı´mica,Escola Te`cnica Superior d’Enginyers Industrials de Barcelona, Universitat Polite`cnica de Catalunya, Diagonal 647, E-08028 Barcelona, Spain; and §Departament de Cie`ncies Fisiolo`giques II, Facultat de Medicina, Campus de Bellvitge, Universitat de Barcelona, L’Hospitalet de Llobregat, E-08907 Barcelona, Spain Communicated by K. E. van Holde, Oregon State University, Corvallis, OR, December 30, 2003 (received for review August 11, 2003) We present evidence that chordate protamines have evolved from point mutations, driven by the selective advantage conferred by histone H1. During the final stages of spermatogenesis, the com- the increased efficiency of highly basic molecules to screen the paction of DNA in many organisms is accomplished by the replace- charge of DNA and thus achieve a more compact chromatin ment of histones with a class of arginine-rich proteins called structure. The relatively quick specialization of histone H1 for protamines. In other organisms, however, condensation of sperm sperm chromatin compaction is not unexpected, because sperm DNA can occur with comparable efficiency in the presence of nuclear basic proteins (SNBPs), like many of the reproductive somatic-type histones or, alternatively, an intermediate class of proteins, are among the most rapidly evolving proteins in the proteins called protamine-like proteins. The idea that the highly animal kingdom (14, 15). specialized sperm chromosomal proteins (protamines) and somatic It has been suggested that protamines did not arise from an chromosomal proteins (histones) could be related dates back al- ancient eukaryotic protein but instead have a retroviral origin most to the discovery of these proteins. -

1 Phylogeny of the Families Pyuridae and Styelidae (Stolidobranchiata
* Manuscript 1 Phylogeny of the families Pyuridae and Styelidae (Stolidobranchiata, Ascidiacea) 2 inferred from mitochondrial and nuclear DNA sequences 3 4 Pérez-Portela Ra, b, Bishop JDDb, Davis ARc, Turon Xd 5 6 a Eco-Ethology Research Unit, Instituto Superior de Psicologia Aplicada (ISPA), Rua 7 Jardim do Tabaco, 34, 1149-041 Lisboa, Portugal 8 9 b Marine Biological Association of United Kingdom, The Laboratory Citadel Hill, PL1 10 2PB, Plymouth, UK, and School of Biological Sciences, University of Plymouth PL4 11 8AA, Plymouth, UK 12 13 c School of Biological Sciences, University of Wollongong, Wollongong NSW 2522 14 Australia 15 16 d Centre d’Estudis Avançats de Blanes (CSIC), Accés a la Cala St. Francesc 14, Blanes, 17 Girona, E-17300, Spain 18 19 Email addresses: 20 Bishop JDD: [email protected] 21 Davis AR: [email protected] 22 Turon X: [email protected] 23 24 Corresponding author: 25 Rocío Pérez-Portela 26 Eco-Ethology Research Unit, Instituto Superior de Psicologia Aplicada (ISPA), Rua 27 Jardim do Tabaco, 34, 1149-041 Lisboa, Portugal 28 Phone: + 351 21 8811226 29 Fax: + 351 21 8860954 30 [email protected] 31 1 32 Abstract 33 34 The Order Stolidobranchiata comprises the families Pyuridae, Styelidae and Molgulidae. 35 Early molecular data was consistent with monophyly of the Stolidobranchiata and also 36 the Molgulidae. Internal phylogeny and relationships between Styelidae and Pyuridae 37 were inconclusive however. In order to clarify these points we used mitochondrial and 38 nuclear sequences from 31 species of Styelidae and 25 of Pyuridae. Phylogenetic trees 39 recovered the Pyuridae as a monophyletic clade, and their genera appeared as 40 monophyletic with the exception of Pyura. -

Tropical Marine Invertebrates CAS BI 569 Phylum Chordata by J
Tropical Marine Invertebrates CAS BI 569 Phylum Chordata by J. R. Finnerty Porifera Ctenophora Cnidaria Deuterostomia Ecdysozoa Lophotrochozoa Chordata Arthropoda Annelida Hemichordata Onychophora Mollusca Echinodermata *Nematoda *Platyhelminthes Acoelomorpha Calcispongia Silicispongiae PROTOSTOMIA Phylum Chordata subPhylum VERTEBRATA subPhylum UROCHORDATA class Ascidaceae—the “sea squirts” class Larvaceae—the larvaceans class Thaliaceae—the salps subPhylum CEPHALOCHORDATA—the “lancelets” Subphylum Subphylum Subphylum Phylum Phylum Vertebrata Urochordata Cephalochordata ECHINODERMATA HEMICHORDATA Phylum CHORDATA Notochord Blastopore -> anus Dorsal hollow nerve cord Radial / equal cleavage Pharyngeal gill slits Coelom forms by enterocoely Endostyle Cross-section through a Chick embryo During Neurulation Subphylum Vertebrata Ordovician Origins—Jawless fishes vertebral column paired appendages post-anal tail Jawed fishes, gnathostomes late Ordovician, early Silurian jaws early Devonian / late Devonian tetrapods / amniotes Permian (299-251) Carboniferous subphylum VERTEBRATA (359-299) gnathostomes (with jaws) 1. Vertebral column Devonian 2. Paired appendages (416-359) 3. Jaws 4. Post anal tail Silurian (444-416) Bony fishes Cartilaginous fishes Paleozoic (543-225) Fishes with jaws & paired fins Ray-finned & Lobe-finned fishes Ordivician Jawless fishes (488-444) Vertebrate origins Jawless fishes phylum CHORDATA Cambrian 1. Dorsal nerve cord (542-488) Non-vert chordates “Cambrian explosion” 2. Notochord Rapid appearance of most 3. Gill slits Animal phyla in fossil record Including Phylum Chordata 4. Endostyle Permian (299-251) 4 3 2 1 1.Pre-mammal-like reptiles 2. Snake-lizard ancestor 3.Crocodile-Bird-Dino ancestor 4. Turtles Carboniferous (359-299) amphibians Early reptiles. Devonian (416-359) Vertebrates invade land “Age of amphibians” Silurian (444-416) Bony fishes AMNIOTES Cartilaginous fishes Paleozoic (543-225) Fishes with jaws & paired fins 1. -

British Pelagic Tunicates, and the Choice of the 21 Species
REVIEWS BULLETIN OF MARINE SCIENCE, 34(1): 175-176, 1984 BRITISHPELAGICTUNICATES.J. H. Fraser. Synopses of the British Fauna (New Series) No. 20. Cambridge University Press. 1982. 57 pp., 23 figures, glossary and index. Soft cover. $12.95. The Synopses of the British Fauna, of which this is No. 20 in the series, are designed as field and laboratory pocketbooks for the use of amateur and profes- sional naturalists. They are intended to bridge the gap between popular field guides and professional treatises and monographs. This slim little volume on pelagic tunicates seems to serve this purpose admirably. Each of the 21 species of Appendicularians and Thaliaceans treated in this work is illustrated with line drawings showing the pertinent characters required for identification. Both aggregate and solitary forms are shown when such occur. Descriptions are brief but adequate and there are notes on distribution and abun- dance where pertinent. A very useful account of anatomical structures, life his- tories, ecology and distribution is given for each major group. There is also a glossary, index and a useful list of references. The introduction serves also to inform the reader on methods of capture, preservation and labeling. Complete keys are given for all levels from class to family, genus and species. The author is to be congratulated on the clarity of the couplet definitions. One may be puzzled, however, by the title, British Pelagic Tunicates, and the choice of the 21 species. Under the Appendicularia Fraser states that only Oi- kopleura dioica and Fritillaria borealis are found in British estuaries and inshore waters while Oikopleura labradoriensis is frequent in areas around the British Isles but is unlikely to be found in inshore waters.