Diversity Partitioning During the Cambrian Radiation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Wright and Cherns Supplementary Data Flat
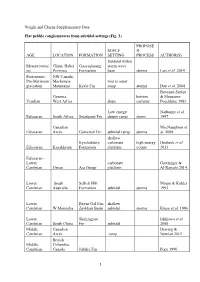
Wright and Cherns Supplementary Data Flat pebble conglomerates from subtidal settings (Fig. 2) PROPOSE SHELF D AGE LOCATION FORMATION SETTING PROCESS AUTHOR(S) Subtidal within Mesoproteroz China, Hebei Gaoyuzhuang storm wave oic Province Formation base storms Luo et al. 2014 Proterozoic, NW Canada, Pre-Marinoan Mackenzie mid to outer glaciation Mountains Keele Fm ramp storms Day et al. 2004 Bertrand-Sarfati Gourma, bottom & Moussine- Vendian West Africa slope currents' Pouchkine 1983 Low energy Narbonne et al. Ediacaran South Africa Swartpunt Fm deeper ramp storm 1997 Canadian MacNaughton et Ediacaran Arctic Gametrail Fm subtidal ramp storms al. 2008 shallow Kyrshabakty carbonate high energy Heubeck et al. Ediacaran Kazakhstan Formation platform events 2013 Ediacaran - Lower carbonate Grotzinger & Cambrian Oman Ara Group platform Al-Rawahi 2014 Lower South Sellick Hill Mount & Kidder Cambrian Australia Formation subtidal storms 1993 Lower Bayan Gol Fm, shallow Cambrian W Mongolia Zavkhan Basin subtidal storms Kruse et al. 1996 Lower Shuijingtuo Ishikawa et al. Cambrian South China Fm subtidal 2008 Middle Canadian Dewing & Cambrian Arctic ramp Nowlan 2012 British Middle Columbia, Cambrian Canada Jubilee Fm Pope 1990 1 Middle Pratt & Cambrian Argentina La Laja Fm subtidal shelf tsunamis Bordonaro 2007 low energy Middle shallow Cambrian Australia Ranken Lst subtidal storms Kruse 1996 Middle Wyoming, Upper Gros Cambrian USA Ventre Shale Csonka 2009 middle upper Middle W Utah, upper Wheeler, carbonate belt - Cambrian USA Marjum fms subtidal shelf Robison 1964 Middle- Upper Supratidal to Cambrian NW China subtidal fpc storms Liang et al. 1993 Ust’- Brus,Labaz, Middle Orakta, Cambrian - Kulyumbe, Lower Ujgur and Iltyk Kouchinsky et Ordovician Siberia fms al. -

Appendix 3.Pdf
A Geoconservation perspective on the trace fossil record associated with the end – Ordovician mass extinction and glaciation in the Welsh Basin Item Type Thesis or dissertation Authors Nicholls, Keith H. Citation Nicholls, K. (2019). A Geoconservation perspective on the trace fossil record associated with the end – Ordovician mass extinction and glaciation in the Welsh Basin. (Doctoral dissertation). University of Chester, United Kingdom. Publisher University of Chester Rights Attribution-NonCommercial-NoDerivatives 4.0 International Download date 26/09/2021 02:37:15 Item License http://creativecommons.org/licenses/by-nc-nd/4.0/ Link to Item http://hdl.handle.net/10034/622234 International Chronostratigraphic Chart v2013/01 Erathem / Era System / Period Quaternary Neogene C e n o z o i c Paleogene Cretaceous M e s o z o i c Jurassic M e s o z o i c Jurassic Triassic Permian Carboniferous P a l Devonian e o z o i c P a l Devonian e o z o i c Silurian Ordovician s a n u a F y r Cambrian a n o i t u l o v E s ' i k s w o Ichnogeneric Diversity k p e 0 10 20 30 40 50 60 70 S 1 3 5 7 9 11 13 15 17 19 21 n 23 r e 25 d 27 o 29 M 31 33 35 37 39 T 41 43 i 45 47 m 49 e 51 53 55 57 59 61 63 65 67 69 71 73 75 77 79 81 83 85 87 89 91 93 Number of Ichnogenera (Treatise Part W) Ichnogeneric Diversity 0 10 20 30 40 50 60 70 1 3 5 7 9 11 13 15 17 19 21 n 23 r e 25 d 27 o 29 M 31 33 35 37 39 T 41 43 i 45 47 m 49 e 51 53 55 57 59 61 c i o 63 z 65 o e 67 a l 69 a 71 P 73 75 77 79 81 83 n 85 a i r 87 b 89 m 91 a 93 C Number of Ichnogenera (Treatise Part W) -

Lee-Riding-2018.Pdf
Earth-Science Reviews 181 (2018) 98–121 Contents lists available at ScienceDirect Earth-Science Reviews journal homepage: www.elsevier.com/locate/earscirev Marine oxygenation, lithistid sponges, and the early history of Paleozoic T skeletal reefs ⁎ Jeong-Hyun Leea, , Robert Ridingb a Department of Geology and Earth Environmental Sciences, Chungnam National University, Daejeon 34134, Republic of Korea b Department of Earth and Planetary Sciences, University of Tennessee, Knoxville, TN 37996, USA ARTICLE INFO ABSTRACT Keywords: Microbial carbonates were major components of early Paleozoic reefs until coral-stromatoporoid-bryozoan reefs Cambrian appeared in the mid-Ordovician. Microbial reefs were augmented by archaeocyath sponges for ~15 Myr in the Reef gap early Cambrian, by lithistid sponges for the remaining ~25 Myr of the Cambrian, and then by lithistid, calathiid Dysoxia and pulchrilaminid sponges for the first ~25 Myr of the Ordovician. The factors responsible for mid–late Hypoxia Cambrian microbial-lithistid sponge reef dominance remain unclear. Although oxygen increase appears to have Lithistid sponge-microbial reef significantly contributed to the early Cambrian ‘Explosion’ of marine animal life, it was followed by a prolonged period dominated by ‘greenhouse’ conditions, as sea-level rose and CO2 increased. The mid–late Cambrian was unusually warm, and these elevated temperatures can be expected to have lowered oxygen solubility, and to have promoted widespread thermal stratification resulting in marine dysoxia and hypoxia. Greenhouse condi- tions would also have stimulated carbonate platform development, locally further limiting shallow-water cir- culation. Low marine oxygenation has been linked to episodic extinctions of phytoplankton, trilobites and other metazoans during the mid–late Cambrian. -

The Ediacaran–Cambrian Transition: the Emerging Record from Small Carbonaceous Fossils (Scfs)
38 The Ediacaran–Cambrian transition: the emerging record from Small Carbonaceous Fossils (SCFs) Ben J. Slater1, Thomas H.P. Harvey2, Romain Guilbaud3, Sebastian Willman1, Graham E. Budd1, Nicholas J. Butterfield4. 1Department of Earth Sciences (Palaeobiology), Uppsala University, Villavägen 16, SE-75236 Uppsala, Sweden ([email protected]) 2School of Geography, Geology and the Environment, University of Leicester, Leicester LE1 7RH, UK 3Lancaster Environment Centre, Lancaster University, Lancaster, LA1 4YQ, UK. 4Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK Abstract.––The most profound change in the fossil record is centered on the Ediacaran–Cambrian transition. Sediments deposited near to this boundary record the first complex trace fossils, the first animal biomineralizers and the earliest fossils of the major animal groups that came to define the subsequent Phanerozoic fossil record. Here we discuss how the emerging record of small carbonaceous fossils (SCFs) is shedding new light on unmineralized aspects of the biota from this critical evolutionary transition. The new SCFs record complements currently established records from trace fossils and biomineralized shelly-remains in reconstructing the broader scale macroevolutionary patterns of this transition, but via an entirely different taphonomic window. SCFs extend the record of multiple major animal groups, improve our picture of protistan diversity from this interval, and reveal new data on elements of the biota that span the Ediacaran–Cambrian boundary. Key words: small carbonaceous fossils; bilateria; Burgess Shale-type preservation; Ediacaran; Terreneuvian. The dramatic changes in the fossil record across subsequent Cambrian radiation (Butterfield, the Ediacaran–Cambrian transition seemingly 2003; Slater et al., 2018a). -

The Weeks Formation Konservat-Lagerstätte and the Evolutionary Transition of Cambrian Marine Life
Downloaded from http://jgs.lyellcollection.org/ by guest on October 1, 2021 Review focus Journal of the Geological Society Published Online First https://doi.org/10.1144/jgs2018-042 The Weeks Formation Konservat-Lagerstätte and the evolutionary transition of Cambrian marine life Rudy Lerosey-Aubril1*, Robert R. Gaines2, Thomas A. Hegna3, Javier Ortega-Hernández4,5, Peter Van Roy6, Carlo Kier7 & Enrico Bonino7 1 Palaeoscience Research Centre, School of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia 2 Geology Department, Pomona College, Claremont, CA 91711, USA 3 Department of Geology, Western Illinois University, 113 Tillman Hall, 1 University Circle, Macomb, IL 61455, USA 4 Department of Zoology, University of Cambridge, Downing Street, Cambridge CB2 3EJ, UK 5 Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, USA 6 Department of Geology, Ghent University, Krijgslaan 281/S8, B-9000 Ghent, Belgium 7 Back to the Past Museum, Carretera Cancún, Puerto Morelos, Quintana Roo 77580, Mexico R.L.-A., 0000-0003-2256-1872; R.R.G., 0000-0002-3713-5764; T.A.H., 0000-0001-9067-8787; J.O.-H., 0000-0002- 6801-7373 * Correspondence: [email protected] Abstract: The Weeks Formation in Utah is the youngest (c. 499 Ma) and least studied Cambrian Lagerstätte of the western USA. It preserves a diverse, exceptionally preserved fauna that inhabited a relatively deep water environment at the offshore margin of a carbonate platform, resembling the setting of the underlying Wheeler and Marjum formations. However, the Weeks fauna differs significantly in composition from the other remarkable biotas of the Cambrian Series 3 of Utah, suggesting a significant Guzhangian faunal restructuring. -

Ediacaran and Cambrian Stratigraphy in Estonia: an Updated Review
Estonian Journal of Earth Sciences, 2017, 66, 3, 152–160 https://doi.org/10.3176/earth.2017.12 Ediacaran and Cambrian stratigraphy in Estonia: an updated review Tõnu Meidla Department of Geology, Institute of Ecology and Earth Sciences, Faculty of Science and Technology, University of Tartu, Ravila 14a, 50411 Tartu, Estonia; [email protected] Received 18 December 2015, accepted 18 May 2017, available online 6 July 2017 Abstract. Previous late Precambrian and Cambrian correlation charts of Estonia, summarizing the regional stratigraphic nomenclature of the 20th century, date back to 1997. The main aim of this review is updating these charts based on recent advances in the global Precambrian and Cambrian stratigraphy and new data from regions adjacent to Estonia. The term ‘Ediacaran’ is introduced for the latest Precambrian succession in Estonia to replace the formerly used ‘Vendian’. Correlation with the dated sections in adjacent areas suggests that only the latest 7–10 Ma of the Ediacaran is represented in the Estonian succession. The gap between the Ediacaran and Cambrian may be rather substantial. The global fourfold subdivision of the Cambrian System is introduced for Estonia. The lower boundary of Series 2 is drawn at the base of the Sõru Formation and the base of Series 3 slightly above the former lower boundary of the ‘Middle Cambrian’ in the Baltic region, marked by a gap in the Estonian succession. The base of the Furongian is located near the base of the Petseri Formation. Key words: Ediacaran, Cambrian, correlation chart, biozonation, regional stratigraphy, Estonia, East European Craton. INTRODUCTION The latest stratigraphic chart of the Cambrian System in Estonia (Mens & Pirrus 1997b, p. -

C0011 Carbon Isotope Stratigraphy
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s), reviewer(s), Elsevier and typesetter TNQ Books and Journals Pvt Ltd. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher and is confidential until formal publication. Chapter 11 c0011 Carbon Isotope Stratigraphy M.R. Saltzman and E. Thomas Chapter Outline 11.1. Principles of Carbon Isotope Stratigraphy 221 11.3.3. Diagenesis 231 11.2. Spatial Heterogeneity of d13C of Dissolved 11.3.4. Global Versus Local Water Mass Signals 232 Inorganic Carbon 223 11.4. Correlation Potential and Excursions 232 11.3. Materials and Methods 224 11.5. Causes of Carbon Isotope Excursions 236 11.3.1. Depositional Setting: Deep (Pelagic) 11.6. Conclusion 237 Versus Shallow 224 Acknowledgments 237 11.3.2. Bulk Versus Component 231 References 237 s0010 11.1. PRINCIPLES OF CARBON ISOTOPE STRATIGRAPHY with the ocean-atmosphere system on longer time scales (105e106 years), contains carbon in both organic matter and p0015 The potential of marine carbonate d13C values to date and carbonate rock (limestones and dolomites) (Berner, 1990). 13 12 correlate rocks relies on the fact that their C/ C values have The carbon isotopic composition of CO2 in the atmo- p0025 varied over time, mainly as the result of partitioning of carbon sphere was about À6.4 & prior to anthropogenic fossil fuel between organic carbon and carbonate carbon reservoirs in the burning, close to the mantle value of about À6& (Sundquist lithosphere (e.g., Shackleton and Hall, 1984; Berner, 1990; and Visser, 2004) (Figure 11.1). -

GSSP) of the Drumian Stage (Cambrian) in the Drum Mountains, Utah, USA
Articles 8585 by Loren E. Babcock1, Richard A. Robison2, Margaret N. Rees3, Shanchi Peng4, and Matthew R. Saltzman1 The Global boundary Stratotype Section and Point (GSSP) of the Drumian Stage (Cambrian) in the Drum Mountains, Utah, USA 1 School of Earth Sciences, The Ohio State University, 125 South Oval Mall, Columbus, OH 43210, USA. Email: [email protected] and [email protected] 2 Department of Geology, University of Kansas, Lawrence, KS 66045, USA. Email: [email protected] 3 Department of Geoscience, University of Nevada, Las Vegas, Las Vegas, NV 89145, USA. Email: [email protected] 4 State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, 39 East Beijing Road, Nanjing 210008, China. Email: [email protected] The Global boundary Stratotype Section and Point correlated with precision through all major Cambrian regions. (GSSP) for the base of the Drumian Stage (Cambrian Among the methods that should be considered in the selection of a GSSP (Remane et al., 1996), biostratigraphic, chemostratigraphic, Series 3) is defined at the base of a limestone (cal- paleogeographic, facies-relationship, and sequence-stratigraphic cisiltite) layer 62 m above the base of the Wheeler For- information is available (e.g., Randolph, 1973; White, 1973; McGee, mation in the Stratotype Ridge section, Drum Moun- 1978; Dommer, 1980; Grannis, 1982; Robison, 1982, 1999; Rowell et al. 1982; Rees 1986; Langenburg et al., 2002a, 2002b; Babcock et tains, Utah, USA. The GSSP level contains the lowest al., 2004; Zhu et al., 2006); that information is summarized here. occurrence of the cosmopolitan agnostoid trilobite Pty- Voting members of the International Subcommission on Cam- chagnostus atavus (base of the P. -

The Auxiliary Boundary Stratotype Section and Point (ASSP) of the Jiangshanian Stage (Cambrian: Furongian Series) in the Kyrshabakty Section
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/261721078 The Auxiliary boundary Stratotype Section and Point (ASSP) of the Jiangshanian Stage (Cambrian: Furongian Series) in the Kyrshabakty section Article in Episodes · March 2014 DOI: 10.18814/epiiugs/2014/v37i1/005 CITATION READS 1 238 7 authors, including: Leonid E. Popov Michael G. Bassett National Museum Wales National Museum of Wales 296 PUBLICATIONS 3,956 CITATIONS 46 PUBLICATIONS 557 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Early Palaeozoic brachiopods and associated faunas of CAOB. View project Cambrian-Ordovician Boundary View project All content following this page was uploaded by Leonid E. Popov on 21 April 2014. The user has requested enhancement of the downloaded file. 41 by Gappar Kh. Ergaliev1, Vyacheslav (Slava) G. Zhemchuzhnikov2, Leonid E. Popov3, Michael G. Bassett3 and Farkhat G. Ergaliev4 The Auxiliary boundary Stratotype Section and Point (ASSP) of the Jiangshanian Stage (Cambrian: Furongian Series) in the Kyrshabakty section, Kazakhstan 1 K.I.Satpaev Institute of Geological Sciences, Kabanbai Batyra Street, 69-a, 050010 Almaty, Kazakhstan. E-mail: [email protected] 2 North Caspian Oil Development LLP, Almaty, Kazakhstan. E-mail: [email protected] 3 Department of Geology, National Museum of Wales, Cathays Park, Cardiff CF10 3NP, Wales, United Kingdom. E-mail: [email protected] 4 Dala Mining (TMM International), Koshek-Batyra Street, 5, 050010 Almaty Kazakhstan. E-mail: [email protected] The Kyrshabakty section in the Malyi Karatau Range, Union of Geological Sciences (IUGS). The GSSP for the stage, now southern Kazakhstan, has been approved as the Auxiliary known as the Jiangshanian (formerly provisional Stage 9) is located in the Duibian B section, Zhejiang, China (Peng et al., 2012). -

(Upper Cambrian, Paibian) Trilobite Faunule in the Central Conasauga River Valley, North Georgia, Usa
Schwimmer.fm Page 31 Monday, June 18, 2012 11:54 AM SOUTHEASTERN GEOLOGY V. 49, No. 1, June 2012, p. 31-41 AN APHELASPIS ZONE (UPPER CAMBRIAN, PAIBIAN) TRILOBITE FAUNULE IN THE CENTRAL CONASAUGA RIVER VALLEY, NORTH GEORGIA, USA DAVID R. SCHWIMMER1 WILLIAM M. MONTANTE2 1Department of Chemistry & Geology Columbus State University, 4225 University Avenue, Columbus, Georgia 31907, USA <[email protected]> 2Marsh & McLennan, Inc., 3560 Lenox Road, Suite 2400, Atlanta, Georgia 30326, USA <[email protected]> ABSTRACT shelf-to-basin break, which is interpreted to be east of the Alabama Promontory and in Middle and Upper Cambrian strata the Tennessee Embayment. The preserva- (Cambrian Series 3 and Furongian) in the tion of abundant aphelaspine specimens by southernmost Appalachians (Tennessee to bioimmuration events may have been the re- Alabama) comprise the Conasauga Forma- sult of mudflows down the shelf-to-basin tion or Group. Heretofore, the youngest re- slope. ported Conasauga beds in the Valley and Ridge Province of Georgia were of the late INTRODUCTION Middle Cambrian (Series 3: Drumian) Bo- laspidella Zone, located on the western state Trilobites and associated biota from Middle boundary in the valley of the Coosa River. Cambrian beds of the Conasauga Formation in Two new localities sited eastward in the Co- northwestern Georgia have been described by nasauga River Valley, yield a diagnostic suite Walcott, 1916a, 1916b; Butts, 1926; Resser, of trilobites from the Upper Cambrian 1938; Palmer, 1962; Schwimmer, 1989; Aphelaspis Zone. Very abundant, Schwimmer and Montante, 2007. These fossils polymeroid trilobites at the primary locality and deposits come from exposures within the are referable to Aphelaspis brachyphasis, valley of the Coosa River, in Floyd County, which is a species known previously in west- Georgia, and adjoining Cherokee County, Ala- ern North America. -

Low-Latitude Origins of the Four Phanerozoic Evolutionary Faunas
bioRxiv preprint doi: https://doi.org/10.1101/866186; this version posted December 6, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Low-Latitude Origins of the Four Phanerozoic Evolutionary Faunas A. Rojas1*, J. Calatayud1, M. Kowalewski2, M. Neuman1, and M. Rosvall1 1Integrated Science Lab, Department of Physics, Umeå University, SE-901 87 Umeå, Sweden 5 2Florida Museum of Natural History, Division of Invertebrate Paleontology, University of Florida, Gainesville, FL 32611, USA *Correspondence to: [email protected] Abstract: Sepkoski’s hypothesis of Three Great Evolutionary Faunas that dominated 10 Phanerozoic oceans represents a foundational concept of macroevolutionary research. However, the hypothesis lacks spatial information and fails to recognize ecosystem changes in Mesozoic oceans. Using a multilayer network representation of fossil occurrences, we demonstrate that Phanerozoic oceans sequentially harbored four evolutionary faunas: Cambrian, Paleozoic, Mesozoic, and Cenozoic. These mega-assemblages all emerged at low latitudes and dispersed 15 out of the tropics. The Paleozoic–Mesozoic transition was abrupt, coincident with the Permian mass extinction, whereas the Mesozoic–Cenozoic transition was protracted, concurrent with gradual ecological shifts posited by the Mesozoic Marine Revolution. These findings support the notion that long-term ecological changes, historical contingencies, and major geological events all have played crucial roles in shaping the evolutionary history of marine animals. 20 One Sentence Summary: Network analysis reveals that Phanerozoic oceans harbored four evolutionary faunas with variable tempo and underlying causes. -

PHANEROZOIC and PRECAMBRIAN CHRONOSTRATIGRAPHY 2016
PHANEROZOIC and PRECAMBRIAN CHRONOSTRATIGRAPHY 2016 Series/ Age Series/ Age Erathem/ System/ Age Epoch Stage/Age Ma Epoch Stage/Age Ma Era Period Ma GSSP/ GSSA GSSP GSSP Eonothem Eon Eonothem Erathem Period Eonothem Period Eon Era System System Eon Erathem Era 237.0 541 Anthropocene * Ladinian Ediacaran Middle 241.5 Neo- 635 Upper Anisian Cryogenian 4.2 ka 246.8 proterozoic 720 Holocene Middle Olenekian Tonian 8.2 ka Triassic Lower 249.8 1000 Lower Mesozoic Induan Stenian 11.8 ka 251.9 Meso- 1200 Upper Changhsingian Ectasian 126 ka Lopingian 254.2 proterozoic 1400 “Ionian” Wuchiapingian Calymmian Quaternary Pleisto- 773 ka 259.8 1600 cene Calabrian Capitanian Statherian 1.80 Guada- 265.1 Proterozoic 1800 Gelasian Wordian Paleo- Orosirian 2.58 lupian 268.8 2050 Piacenzian Roadian proterozoic Rhyacian Pliocene 3.60 272.3 2300 Zanclean Kungurian Siderian 5.33 Permian 282.0 2500 Messinian Artinskian Neo- 7.25 Cisuralian 290.1 Tortonian Sakmarian archean 11.63 295.0 2800 Serravallian Asselian Meso- Miocene 13.82 298.9 r e c a m b i n P Neogene Langhian Gzhelian archean 15.97 Upper 303.4 3200 Burdigalian Kasimovian Paleo- C e n o z i c 20.44 306.7 Archean archean Aquitanian Penn- Middle Moscovian 23.03 sylvanian 314.6 3600 Chattian Lower Bashkirian Oligocene 28.1 323.2 Eoarchean Rupelian Upper Serpukhovian 33.9 330.9 4000 Priabonian Middle Visean 38.0 Carboniferous 346.7 Hadean (informal) Missis- Bartonian sippian Lower Tournaisian Eocene 41.0 358.9 ~4560 Lutetian Famennian 47.8 Upper 372.2 Ypresian Frasnian Units of the international Paleogene 56.0 382.7 Thanetian Givetian chronostratigraphic scale with 59.2 Middle 387.7 Paleocene Selandian Eifelian estimated numerical ages.