A Quick Look at the Jurassic World
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mesozoic—Dinos!
MESOZOIC—DINOS! VOLUME 9, ISSUE 8, APRIL 2020 THIS MONTH DINOSAURS! • Dinosaurs ○ What is a Dinosaur? page 2 DINOSAURS! When people think paleontology, ○ Bird / Lizard Hip? page 5 they think of scientists ○ Size Activity 1 page 10 working in the hot sun of ○ Size Activity 2 page 13 Colorado National ○ Size Activity 3 page 43 Monument or the Badlands ○ Diet page 46 of South Dakota and ○ Trackways page 53 Wyoming finding enormous, ○ Colorado Fossils and fierce, and long-gone Dinosaurs page 66 dinosaurs. POWER WORDS Dinosaurs safely evoke • articulated: fossil terror. Better than any bones arranged in scary movie, these were Articulated skeleton of the Tyrannosaurus rex proper order actually living breathing • endothermic: an beasts! from the American Museum of Natural History organism produces body heat through What was the biggest dinosaur? be reviewing the information metabolism What was the smallest about dinosaurs, but there is an • metabolism: chemical dinosaur? What color were interview with him at the end of processes that occur they? Did they live in herds? this issue. Meeting him, you will within a living organism What can their skeletons tell us? know instantly that he loves his in order to maintain life What evidence is there so that job! It doesn’t matter if you we can understand more about become an electrician, auto CAREER CONNECTION how these animals lived. Are mechanic, dancer, computer • Meet Dr. Holtz, any still alive today? programmer, author, or Dinosaur paleontologist, I truly hope that Paleontologist! page 73 To help us really understand you have tremendous job more about dinosaurs, we have satisfaction, like Dr. -

Diversity Partitioning During the Cambrian Radiation
Diversity partitioning during the Cambrian radiation Lin Naa,1 and Wolfgang Kiesslinga,b aGeoZentrum Nordbayern, Paleobiology and Paleoenvironments, Friedrich-Alexander-Universität Erlangen-Nürnberg, 91054 Erlangen, Germany; and bMuseum für Naturkunde, Leibniz Institute for Research on Evolution and Biodiversity at the Humboldt University Berlin, 10115 Berlin, Germany Edited by Douglas H. Erwin, Smithsonian National Museum of Natural History, Washington, DC, and accepted by the Editorial Board March 10, 2015 (received for review January 2, 2015) The fossil record offers unique insights into the environmental and Results geographic partitioning of biodiversity during global diversifica- Raw gamma diversity exhibits a strong increase in the first three tions. We explored biodiversity patterns during the Cambrian Cambrian stages (informally referred to as early Cambrian in this radiation, the most dramatic radiation in Earth history. We as- work) (Fig. 1A). Gamma diversity dropped in Stage 4 and de- sessed how the overall increase in global diversity was partitioned clined further through the rest of the Cambrian. The pattern is between within-community (alpha) and between-community (beta) robust to sampling standardization (Fig. 1B) and insensitive to components and how beta diversity was partitioned among environ- including or excluding the archaeocyath sponges, which are po- ments and geographic regions. Changes in gamma diversity in the tentially oversplit (16). Alpha and beta diversity increased from Cambrian were chiefly driven by changes in beta diversity. The the Fortunian to Stage 3, and fluctuated erratically through the combined trajectories of alpha and beta diversity during the initial following stages (Fig. 2). Our estimate of alpha (and indirectly diversification suggest low competition and high predation within beta) diversity is based on the number of genera in published communities. -

Triassic- Jurassic Stratigraphy Of
Triassic- Jurassic Stratigraphy of the <JF C7 JL / Culpfeper and B arbour sville Basins, VirginiaC7 and Maryland/ ll.S. PAPER Triassic-Jurassic Stratigraphy of the Culpeper and Barboursville Basins, Virginia and Maryland By K.Y. LEE and AJ. FROELICH U.S. GEOLOGICAL SURVEY PROFESSIONAL PAPER 1472 A clarification of the Triassic--Jurassic stratigraphic sequences, sedimentation, and depositional environments UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON: 1989 DEPARTMENT OF THE INTERIOR MANUEL LUJAN, Jr., Secretary U.S. GEOLOGICAL SURVEY Dallas L. Peck, Director Any use of trade, product, or firm names in this publication is for descriptive purposes only and does not imply endorsement by the U.S. Government Library of Congress Cataloging in Publication Data Lee, K.Y. Triassic-Jurassic stratigraphy of the Culpeper and Barboursville basins, Virginia and Maryland. (U.S. Geological Survey professional paper ; 1472) Bibliography: p. Supt. of Docs. no. : I 19.16:1472 1. Geology, Stratigraphic Triassic. 2. Geology, Stratigraphic Jurassic. 3. Geology Culpeper Basin (Va. and Md.) 4. Geology Virginia Barboursville Basin. I. Froelich, A.J. (Albert Joseph), 1929- II. Title. III. Series. QE676.L44 1989 551.7'62'09755 87-600318 For sale by the Books and Open-File Reports Section, U.S. Geological Survey, Federal Center, Box 25425, Denver, CO 80225 CONTENTS Page Page Abstract.......................................................................................................... 1 Stratigraphy Continued Introduction... .......................................................................................... -

The Jurassic Pleurotomarioidean Gastropod Laevitomaria and Its Palaeobiogeographical History
The Jurassic pleurotomarioidean gastropod Laevitomaria and its palaeobiogeographical history ROBERTO GATTO, STEFANO MONARI, JÁNOS SZABÓ, and MARIA ALESSANDRA CONTI Gatto, R., Monari, S., Szabó, J., and Conti, M.A. 2015. The Jurassic pleurotomarioidean gastropod Laevitomaria and its palaeobiogeographical history. Acta Palaeontologica Polonica 60 (1): 217–233. The genus Laevitomaria is reviewed and its palaeobiogeographical history is reconstructed based on the re-examination of its type species L. problematica, the study of material stored at the National Natural History Museum of Luxembourg, and an extensive review of the literature. The systematic study allows ascribing to Laevitomaria a number of Jurassic species from the western European region formerly included in other pleurotomariid genera. The following new combi- nations are proposed: Laevitomaria allionta, L. amyntas, L. angulba, L. asurai, L. daityai, L. fasciata, L. gyroplata, L. isarensis, L. joannis, L. repeliniana, L. stoddarti, L. subplatyspira, and L. zonata. The genus, which was once considered as endemic of the central part of the western Tethys, shows an evolutionary and palaeogeographical history consider- ably more complex than previously assumed. It first appeared in the Late Sinemurian in the northern belt of the central western Tethys involved in the Neotethyan rifting, where it experienced a first radiation followed by an abrupt decline of diversity in the Toarcian. Species diversity increased again during Toarcian–Aalenian times in the southernmost part of western European shelf and a major radiation occurred during the Middle Aalenian to Early Bajocian in the northern Paris Basin and southern England. After a latest Bajocian collapse of diversity, Laevitomaria disappeared from both the central part of western Tethys and the European shelf. -

Thomas Jefferson Meg Tooth
The ECPHORA The Newsletter of the Calvert Marine Museum Fossil Club Volume 30 Number 3 September 2015 Thomas Jefferson Meg Tooth Features Thomas Jefferson Meg The catalogue number Review; Walking is: ANSP 959 Whales Inside The tooth came from Ricehope Estate, Snaggletooth Shark Cooper River, Exhibit South Carolina. Tiktaalik Clavatulidae In 1806, it was Juvenile Bald Eagle originally collected or Sculpting Whale Shark owned by Dr. William Moroccan Fossils Reid. Prints in the Sahara Volunteer Outing to Miocene-Pliocene National Geographic coastal plain sediments. Dolphins in the Chesapeake Sloth Tooth Found SharkFest Shark Iconography in Pre-Columbian Panama Hippo Skulls CT- Scanned Squalus sp. Teeth Sperm Whale Teeth On a recent trip to the Academy of Natural Sciences of Drexel University (Philadelphia), Collections Manager Ned Gilmore gave John Nance and me a behind -the-scenes highlights tour. Among the fossils that belonged to Thomas☼ Jefferson (left; American Founding Father, principal author of the Declaration of Independence, and third President of the United States) was this Carcharocles megalodon tooth. Jefferson’s interests and knowledge were encyclopedic; a delight to know that they included paleontology. Hand by J. Nance. Photo by S. Godfrey. Jefferson portrait from: http://www.biography.com/people/thomas-jefferson-9353715 ☼ CALVERT MARINE MUSEUM www.calvertmarinemuseum.com 2 The Ecphora September 2015 Book Review: The Walking 41 million years ago and has worldwide distribution. It was fully aquatic, although it did have residual Whales hind limbs. In later chapters, Professor Thewissen George F. Klein discusses limb development and various genetic factors that make whales, whales. This is a The full title of this book is The Walking complicated topic, but I found these chapters very Whales — From Land to Water in Eight Million clear and readable. -

The Geologic Time Scale Is the Eon
Exploring Geologic Time Poster Illustrated Teacher's Guide #35-1145 Paper #35-1146 Laminated Background Geologic Time Scale Basics The history of the Earth covers a vast expanse of time, so scientists divide it into smaller sections that are associ- ated with particular events that have occurred in the past.The approximate time range of each time span is shown on the poster.The largest time span of the geologic time scale is the eon. It is an indefinitely long period of time that contains at least two eras. Geologic time is divided into two eons.The more ancient eon is called the Precambrian, and the more recent is the Phanerozoic. Each eon is subdivided into smaller spans called eras.The Precambrian eon is divided from most ancient into the Hadean era, Archean era, and Proterozoic era. See Figure 1. Precambrian Eon Proterozoic Era 2500 - 550 million years ago Archaean Era 3800 - 2500 million years ago Hadean Era 4600 - 3800 million years ago Figure 1. Eras of the Precambrian Eon Single-celled and simple multicelled organisms first developed during the Precambrian eon. There are many fos- sils from this time because the sea-dwelling creatures were trapped in sediments and preserved. The Phanerozoic eon is subdivided into three eras – the Paleozoic era, Mesozoic era, and Cenozoic era. An era is often divided into several smaller time spans called periods. For example, the Paleozoic era is divided into the Cambrian, Ordovician, Silurian, Devonian, Carboniferous,and Permian periods. Paleozoic Era Permian Period 300 - 250 million years ago Carboniferous Period 350 - 300 million years ago Devonian Period 400 - 350 million years ago Silurian Period 450 - 400 million years ago Ordovician Period 500 - 450 million years ago Cambrian Period 550 - 500 million years ago Figure 2. -

The Cenozoic Era - Nýlífsöld 65 MY-Present Jarðsaga 2 Ólafur Ingólfsson Origin of the Term: the Tertiary Tertiary System
The Cenozoic Era - Nýlífsöld 65 MY-Present Jarðsaga 2 Ólafur Ingólfsson Origin of the Term: The Tertiary Tertiary System. [1760] Named by Giovanni Arduino Period as the uppermost part of his 65-1.8 MY three-fold subdivision of mountains in northern Italy. The Tertiary became a formal period and system when Lyell published his work describing further subdivisions of the Tertiary. The Tertiary Period is divided into five epochs (tímar): Paleocene (65-56 MY), Eocene (56-34 MY), Oligocene (34-24 MY), Miocene (24-5,3 MY), and Pliocene (5,3-1,8 MY). Confusing set of stratigraphic terms... More than 95% of the Cenozoic era belongs to the Tertiary period. During the 18th century the names Primary, Secondary, and Tertiary were given by Giovanni Arduino to successive rock strata, the Primary being the oldest, the Tertiary the more recent. In 1829 a fourth division, the Quaternary, was added by P. G. Desnoyers. These terms were later abandoned, the Primary becoming the Paleozoic Era, and the Secondary the Mesozoic. But Tertiary and Quaternary were retained for the two main stages of the Cenozoic. Attempts to replace the "Tertiary" with a more reasonable division of “Palaeogene” (early Tertiary) and “Neogene” (later Tertiary and Quaternary) have not been very successful. Stanley uses this division. The World at the K/T Boundary Paleocene plate tectonics During the Paleocene, the inland seas of the Cretaceous Period dry up, exposing large land areas in North America and Eurasia. Australia begins to separate from Antarctica, and Greenland splits from North America. A remnant Tethys Sea persists in the equatorial region. -

Recent Advances in Studies on Mesozoic and Paleogene Mammals in China
Vol.24 No.2 2010 Paleomammalogy Recent Advances in Studies on Mesozoic and Paleogene Mammals in China WANG Yuanqing* and NI Xijun Institute of Vertebrate Paleontology and Paleoanthropology, CAS, Beijing 10004, China ike in other fields of paleontology, research in from the articular of the lower jaw and the quadrate of the paleomammalogy mainly falls into two aspects. cranium following the process of reduction and detachment LOne is related to the biological nature of fossil from the reptilian mandible, are new elements of the bony mammals, such as their systematics, origin, evolution, chain in the mammalian middle ear. The appearance of phylogenetic relationships, transformation of key features mammalian middle ear allows mammals to hear the sound and paleobiogeography, and the other is related to the of higher frequencies than reptiles do. Generally speaking, geological context, involving biostratigraphy, biochronology, a widely accepted hypothesis is that mammals originated faunal turnover and its relations to the global and regional from a certain extinct reptilian group. The formation and environmental changes. In the last decade, a number of development of the definitive mammalian middle ear well-preserved mammalian specimens were collected from (DMME) has thus become one of the key issues in the study different localities around the country. Such discoveries have of mammalian evolution and has drawn the attention from provided significant information for understanding the early many researchers for many years. evolution of mammals and reconstructing the phylogeny of Developmental biological studies have proven the early mammals. function of the Meckel’s cartilage and its relationship to Studies of Chinese Mesozoic mammals achieved the ear ossicles during the development of mammalian remarkable progress in the past several years. -
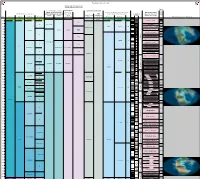
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

Crustacea: Thalassinidea, Brachyura) from Puerto Rico, United States Territory
Bulletin of the Mizunami Fossil Museum, no. 34 (2008), p. 1–15, 6 figs., 1 table. © 2008, Mizunami Fossil Museum New Cretaceous and Cenozoic Decapoda (Crustacea: Thalassinidea, Brachyura) from Puerto Rico, United States Territory Carrie E. Schweitzer1, Jorge Velez-Juarbe2, Michael Martinez3, Angela Collmar Hull1, 4, Rodney M. Feldmann4, and Hernan Santos2 1)Department of Geology, Kent State University Stark Campus, 6000 Frank Ave. NW, North Canton, Ohio, 44720, USA <[email protected]> 2)Department of Geology, University of Puerto Rico, Mayagüez Campus, P. O. Box 9017, Mayagüez, Puerto Rico, 00681 United States Territory <[email protected]> 3)College of Marine Science, University of South Florida, 140 7th Ave. South, St. Petersburg, Florida 33701, USA <[email protected]> 4)Department of Geology, Kent State University, Kent, Ohio 44242, USA <[email protected]> Abstract A large number of recently collected specimens from Puerto Rico has yielded two new species including Palaeoxanthopsis tylotus and Eurytium granulosum, the oldest known occurrence of the latter genus. Cretaceous decapods are reported from Puerto Rico for the first time, and the Cretaceous fauna is similar to that of southern Mexico. Herein is included the first report of Pleistocene decapods from Puerto Rico, which were previously known from other Caribbean localities. The Pleistocene Cardisoma guanhumi is a freshwater crab of the family Gecarcinidae. The freshwater crab families have a poor fossil record; thus, the occurrence is noteworthy and may document dispersal of the crab by humans. Key words: Decapoda, Thalassinidea, Brachyura, Puerto Rico, Cretaceous, Paleogene, Neogene. Introduction than Eocene are not separated by these fault zones and even overlie parts of the fault zones in some areas (Jolly et al., 1998). -

The Late Jurassic Tithonian, a Greenhouse Phase in the Middle Jurassic–Early Cretaceous ‘Cool’ Mode: Evidence from the Cyclic Adriatic Platform, Croatia
Sedimentology (2007) 54, 317–337 doi: 10.1111/j.1365-3091.2006.00837.x The Late Jurassic Tithonian, a greenhouse phase in the Middle Jurassic–Early Cretaceous ‘cool’ mode: evidence from the cyclic Adriatic Platform, Croatia ANTUN HUSINEC* and J. FRED READ *Croatian Geological Survey, Sachsova 2, HR-10000 Zagreb, Croatia Department of Geosciences, Virginia Tech, 4044 Derring Hall, Blacksburg, VA 24061, USA (E-mail: [email protected]) ABSTRACT Well-exposed Mesozoic sections of the Bahama-like Adriatic Platform along the Dalmatian coast (southern Croatia) reveal the detailed stacking patterns of cyclic facies within the rapidly subsiding Late Jurassic (Tithonian) shallow platform-interior (over 750 m thick, ca 5–6 Myr duration). Facies within parasequences include dasyclad-oncoid mudstone-wackestone-floatstone and skeletal-peloid wackestone-packstone (shallow lagoon), intraclast-peloid packstone and grainstone (shoal), radial-ooid grainstone (hypersaline shallow subtidal/intertidal shoals and ponds), lime mudstone (restricted lagoon), fenestral carbonates and microbial laminites (tidal flat). Parasequences in the overall transgressive Lower Tithonian sections are 1– 4Æ5 m thick, and dominated by subtidal facies, some of which are capped by very shallow-water grainstone-packstone or restricted lime mudstone; laminated tidal caps become common only towards the interior of the platform. Parasequences in the regressive Upper Tithonian are dominated by peritidal facies with distinctive basal oolite units and well-developed laminate caps. Maximum water depths of facies within parasequences (estimated from stratigraphic distance of the facies to the base of the tidal flat units capping parasequences) were generally <4 m, and facies show strongly overlapping depth ranges suggesting facies mosaics. Parasequences were formed by precessional (20 kyr) orbital forcing and form parasequence sets of 100 and 400 kyr eccentricity bundles. -

A Comparison of the Dinosaur Communities from the Middle
Preprints (www.preprints.org) | NOT PEER-REVIEWED | Posted: 31 July 2018 doi:10.20944/preprints201807.0610.v1 Peer-reviewed version available at Geosciences 2018, 8, 327; doi:10.3390/geosciences8090327 1 Review 2 A comparison of the dinosaur communities from 3 the Middle Jurassic of the Cleveland (Yorkshire) 4 and Hebrides (Skye) basins, based on their ichnites 5 6 Mike Romano 1*, Neil D. L. Clark 2 and Stephen L. Brusatte 3 7 1 Independent Researcher, 14 Green Lane, Dronfield, Sheffield S18 2LZ, England, United Kingdom; 8 [email protected] 9 2 Curator of Palaeontology, The Hunterian, University of Glasgow, University Avenue, Glasgow 10 G12 8QQ, Scotland, United Kingdom; [email protected] 11 3 Chancellor's Fellow in Vertebrate Palaeontology, School of Geosciences, University of Edinburgh, 12 Grant Institute, The King's Buildings, James Hutton Road, Edinburgh EH9 3FE, Scotland, United Kingdom; 13 [email protected] 14 * Correspondence: [email protected]; Tel.: 01246 417330 15 16 Abstract: 17 Despite the Hebrides and Cleveland basins being geographically close, research has not 18 previously been carried out to determine faunal similarities and assess the possibility of links 19 between the dinosaur populations. The palaeogeography of both areas during the Middle Jurassic 20 shows that there were no elevated landmasses being eroded to produce conglomeratic material in 21 the basins at that time. The low-lying landscape and connected shorelines may have provided 22 connectivity between the two dinosaur populations. 23 The dinosaur fauna of the Hebrides and Cleveland basins has been assessed based primarily 24 on the abundant ichnites found in both areas as well as their skeletal remains.