The Ecology and Ecological Enhancement of Artificial Coastal Structures
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Management of Coastal Erosion by Creating Large-Scale and Small-Scale Sediment Cells
COASTAL EROSION CONTROL BASED ON THE CONCEPT OF SEDIMENT CELLS by L. C. van Rijn, www.leovanrijn-sediment.com, March 2013 1. Introduction Nearly all coastal states have to deal with the problem of coastal erosion. Coastal erosion and accretion has always existed and these processes have contributed to the shaping of the present coastlines. However, coastal erosion now is largely intensified due to human activities. Presently, the total coastal area (including houses and buildings) lost in Europe due to marine erosion is estimated to be about 15 km2 per year. The annual cost of mitigation measures is estimated to be about 3 billion euros per year (EUROSION Study, European Commission, 2004), which is not acceptable. Although engineering projects are aimed at solving the erosion problems, it has long been known that these projects can also contribute to creating problems at other nearby locations (side effects). Dramatic examples of side effects are presented by Douglas et al. (The amount of sand removed from America’s beaches by engineering works, Coastal Sediments, 2003), who state that about 1 billion m3 (109 m3) of sand are removed from the beaches of America by engineering works during the past century. The EUROSION study (2004) recommends to deal with coastal erosion by restoring the overall sediment balance on the scale of coastal cells, which are defined as coastal compartments containing the complete cycle of erosion, deposition, sediment sources and sinks and the transport paths involved. Each cell should have sufficient sediment reservoirs (sources of sediment) in the form of buffer zones between the land and the sea and sediment stocks in the nearshore and offshore coastal zones to compensate by natural or artificial processes (nourishment) for sea level rise effects and human-induced erosional effects leading to an overall favourable sediment status. -

Animal Adaptations
Animal Adaptations The Animal Adaptations program at Hatfield Marine Science Center is designed to be a 50- minute lab-based program for 3rd-12th grade students that examines marine organisms from three different habitats (sandy beach, rocky shore and estuary) and explores the many ways they are adapted to their particular environment. This lab focuses on the adaptations of several groups of marine animals including Mollusks, Crustaceans, and Echinoderms, and investigates how they differ depending on whether they are found in a sandy beach, rocky shore, or estuary environment. Students will work in small groups with a variety of live animals, studying individual characteristics and how these organisms interact with their environment and one another. Background Information The Oregon Coast is made up of a series of rocky shores, sandy beaches, and estuaries, all of which are greatly affected by fluctuating tides. Many of these areas are intertidal and are alternately inundated by seawater and exposed to air, wind, and dramatic changes in temperature and salinity. High tide floods these areas with cold, nutrient laden seawater, bringing food to organisms that live there in the form of plankton and detritus. Low tides often expose these organisms to the dangers of predation and desiccation. In addition to tidal effects, organisms that inhabit sandy beaches and rocky shores also have to deal with the physical stresses of pounding waves. Because of these harsh conditions, organisms have developed special adaptations that not only help them to survive but thrive in these environments. An adaptation is a physical or behavioral trait that helps a plant or animal survive in a specific environment or habitat. -

Seashore the SANDYSEASHORE the Soilisinfertile, Anditisoften Windy, Andsalty
SEASHORE DESCRIPTION The seashore is an area fi lled with an interesting mix of unique plants and animals that have adapted to cope with this environment. Living on the edge of the sea is not easy. The soil is infertile, and it is often windy, dry and salty. THE SANDY SEASHORE Along a sandy shore there are no large rocks, algae or tidal pools. The sandy seashore can be divided into four general zones: Intertidal, Pioneer, Fixed Dune and Scrub Woodland. 1. Intertidal Zone: Between the low tide and the high tide mark is the intertidal zone. When the tide goes out the creatures living in this zone are left stranded. They have to endure the heat of the sun and the higher salinity of the water resulting from evaporation. Notice the many small holes on a sandy beach; they are the doorways to the homes of many animals which burrow under the sand where it is cooler. Some of Ecosystems of The Bahamas the creatures living in this zone are sea worms, sand fl eas and sand crabs. 2. Pioneer Zone: So named because it is where the fi rst plants to try to grow over sand. These plants must adapt to loose, shifting sand and poor soil. There is no protection from wind or salt spray. Plants here are usually low growing vines with waxy leaves. Some plants found in the pioneer zone are Purple seaside bean (C. rosea), Saltwort (Batis maritima), Goat's foot (Ipomea pes-caprae), and Sea purslane (Sesuvium portulacastrum). 3. Fixed Dune Zone: The next zone is the fi xed dune, so named because as the plants in the pioneer zone grip the sand around their roots and make the beach more stable, the sand mounds up into small humps or dunes. -

1 the Influence of Groyne Fields and Other Hard Defences on the Shoreline Configuration
1 The Influence of Groyne Fields and Other Hard Defences on the Shoreline Configuration 2 of Soft Cliff Coastlines 3 4 Sally Brown1*, Max Barton1, Robert J Nicholls1 5 6 1. Faculty of Engineering and the Environment, University of Southampton, 7 University Road, Highfield, Southampton, UK. S017 1BJ. 8 9 * Sally Brown ([email protected], Telephone: +44(0)2380 594796). 10 11 Abstract: Building defences, such as groynes, on eroding soft cliff coastlines alters the 12 sediment budget, changing the shoreline configuration adjacent to defences. On the 13 down-drift side, the coastline is set-back. This is often believed to be caused by increased 14 erosion via the ‘terminal groyne effect’, resulting in rapid land loss. This paper examines 15 whether the terminal groyne effect always occurs down-drift post defence construction 16 (i.e. whether or not the retreat rate increases down-drift) through case study analysis. 17 18 Nine cases were analysed at Holderness and Christchurch Bay, England. Seven out of 19 nine sites experienced an increase in down-drift retreat rates. For the two remaining sites, 20 retreat rates remained constant after construction, probably as a sediment deficit already 21 existed prior to construction or as sediment movement was restricted further down-drift. 22 For these two sites, a set-back still evolved, leading to the erroneous perception that a 23 terminal groyne effect had developed. Additionally, seven of the nine sites developed a 24 set back up-drift of the initial groyne, leading to the defended sections of coast acting as 1 25 a hard headland, inhabiting long-shore drift. -

PROTISTS Shore and the Waves Are Large, Often the Largest of a Storm Event, and with a Long Period
(seas), and these waves can mobilize boulders. During this phase of the storm the rapid changes in current direction caused by these large, short-period waves generate high accelerative forces, and it is these forces that ultimately can move even large boulders. Traditionally, most rocky-intertidal ecological stud- ies have been conducted on rocky platforms where the substrate is composed of stable basement rock. Projec- tiles tend to be uncommon in these types of habitats, and damage from projectiles is usually light. Perhaps for this reason the role of projectiles in intertidal ecology has received little attention. Boulder-fi eld intertidal zones are as common as, if not more common than, rock plat- forms. In boulder fi elds, projectiles are abundant, and the evidence of damage due to projectiles is obvious. Here projectiles may be one of the most important defi ning physical forces in the habitat. SEE ALSO THE FOLLOWING ARTICLES Geology, Coastal / Habitat Alteration / Hydrodynamic Forces / Wave Exposure FURTHER READING Carstens. T. 1968. Wave forces on boundaries and submerged bodies. Sarsia FIGURE 6 The intertidal zone on the north side of Cape Blanco, 34: 37–60. Oregon. The large, smooth boulders are made of serpentine, while Dayton, P. K. 1971. Competition, disturbance, and community organi- the surrounding rock from which the intertidal platform is formed zation: the provision and subsequent utilization of space in a rocky is sandstone. The smooth boulders are from a source outside the intertidal community. Ecological Monographs 45: 137–159. intertidal zone and were carried into the intertidal zone by waves. Levin, S. A., and R. -

Baja California Sur, Mexico)
Journal of Marine Science and Engineering Article Geomorphology of a Holocene Hurricane Deposit Eroded from Rhyolite Sea Cliffs on Ensenada Almeja (Baja California Sur, Mexico) Markes E. Johnson 1,* , Rigoberto Guardado-France 2, Erlend M. Johnson 3 and Jorge Ledesma-Vázquez 2 1 Geosciences Department, Williams College, Williamstown, MA 01267, USA 2 Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada 22800, Baja California, Mexico; [email protected] (R.G.-F.); [email protected] (J.L.-V.) 3 Anthropology Department, Tulane University, New Orleans, LA 70018, USA; [email protected] * Correspondence: [email protected]; Tel.: +1-413-597-2329 Received: 22 May 2019; Accepted: 20 June 2019; Published: 22 June 2019 Abstract: This work advances research on the role of hurricanes in degrading the rocky coastline within Mexico’s Gulf of California, most commonly formed by widespread igneous rocks. Under evaluation is a distinct coastal boulder bed (CBB) derived from banded rhyolite with boulders arrayed in a partial-ring configuration against one side of the headland on Ensenada Almeja (Clam Bay) north of Loreto. Preconditions related to the thickness of rhyolite flows and vertical fissures that intersect the flows at right angles along with the specific gravity of banded rhyolite delimit the size, shape and weight of boulders in the Almeja CBB. Mathematical formulae are applied to calculate the wave height generated by storm surge impacting the headland. The average weight of the 25 largest boulders from a transect nearest the bedrock source amounts to 1200 kg but only 30% of the sample is estimated to exceed a full metric ton in weight. -

Marine Biological Investigations in the Bahamas 23. Description of the Littoral Zonation at Nine Bahamian Rocky-Shore Localities
Marine biological investigations in the Bahamas 23. Description of the littoral zonation at nine Bahamian rocky-shore localities Hans Brattström Brattström H. 1999. Description of the littoral zonation at nine Bahamian rocky-shore localities. SARSIA Sarsia 84:319-365. Nine typical, more or less vertical, rocky-shore localities were visited in April-May 1967, and one of these was revisited in March 1968. The topographies of the stations are described and illustrated, and the vertical zonation of their flora and fauna is documented. On the whole the flora and fauna at the stations investigated was fairly poor. At least 37 species of algae and 85 animal species were recorded, but only 15 algae and 71 animals could be identified to species. Most of the algae were too undeveloped or grazed down to permit a reliable identification. Only half as many species were found at these nine stations as were found on three almost horizontal beach-rock stations described in an earlier article. Whereas many of the latter lived in a system of cavities below the rocks, all species found on the nine vertical stations were exposed to waves and the hot midday sun. The common yellow, black, grey, and white zones typical of the Bahamas, were often well-devel- oped. Some algae and many of the animals formed distinct belts within these zones, the higher up the more exposed the rocks were. Though tides and exposure presumably are responsible for the general zonation pattern, it is likely that in the cirripeds and vermetids the way of food-intake determines their upper limit. -

Guidance for Flood Risk Analysis and Mapping
Guidance for Flood Risk Analysis and Mapping Coastal Notations, Acronyms, and Glossary of Terms May 2016 Requirements for the Federal Emergency Management Agency (FEMA) Risk Mapping, Assessment, and Planning (Risk MAP) Program are specified separately by statute, regulation, or FEMA policy (primarily the Standards for Flood Risk Analysis and Mapping). This document provides guidance to support the requirements and recommends approaches for effective and efficient implementation. Alternate approaches that comply with all requirements are acceptable. For more information, please visit the FEMA Guidelines and Standards for Flood Risk Analysis and Mapping webpage (www.fema.gov/guidelines-and-standards-flood-risk-analysis-and- mapping). Copies of the Standards for Flood Risk Analysis and Mapping policy, related guidance, technical references, and other information about the guidelines and standards development process are all available here. You can also search directly by document title at www.fema.gov/library. Coastal Notation, Acronyms, and Glossary of Terms May 2016 Guidance Document 66 Page i Document History Affected Section or Date Description Subsection Initial version of new transformed guidance. The content was derived from the Guidelines and Specifications for Flood Hazard Mapping Partners, Procedure Memoranda, First Publication May 2016 and/or Operating Guidance documents. It has been reorganized and is being published separately from the standards. Coastal Notation, Acronyms, and Glossary of Terms May 2016 Guidance Document 66 -
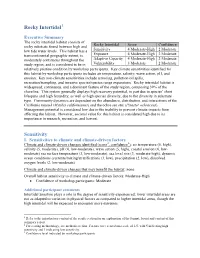
Rocky Intertidal Sensitivity
Rocky Intertidal1 Executive Summary The rocky intertidal habitat consists of Rocky Intertidal Score Confidence rocky substrate found between high and low tide water levels. This habitat has a Sensitivity 4 Moderate-High 2 Moderate transcontinental geographic extent, is Exposure 4 Moderate-High 2 Moderate moderately continuous throughout the Adaptive Capacity 4 Moderate-High 2 Moderate study region, and is considered to be in Vulnerability 3 Moderate 2 Moderate relatively pristine condition by workshop participants. Key climate sensitivities identified for this habitat by workshop participants includes air temperature, salinity, wave action, pH, and erosion. Key non-climate sensitivities include armoring, pollution/oil spills, recreation/trampling, and invasive species/species range expansions. Rocky intertidal habitat is widespread, continuous, and a dominant feature of the study region, composing 39% of the shoreline. This system generally displays high recovery potential, in part due to species’ short lifespans and high fecundity, as well as high species diversity, due to the diversity in substrate type. Community dynamics are dependent on the abundance, distribution, and interactions of the California mussel (Mytilus californianus) and the ochre sea star (Pisaster ochraceus). Management potential is considered low due to the inability to prevent climate impacts from affecting the habitat. However, societal value for this habitat is considered high due to its importance in research, recreation, and harvest. Sensitivity I. Sensitivities -

Mathematical Model of Groynes on Shingle Beaches
HR Wallingford Mathematical Model of Groynes on Shingle Beaches A H Brampton BSc PhD D G Goldberg BA Report SR 276 November 1991 Address:Hydraulics Research Ltd, wallingford,oxfordshire oxl0 gBA,United Kingdom. Telephone:0491 35381 Intemarional + 44 49135381 relex: g4gsszHRSwALG. Facstunile:049132233Intemarional + M 49132233 Registeredin EngtandNo. 1622174 This report describes an investigation carried out by HR Wallingford under contract CSA 1437, 'rMathematical- Model of Groynes on Shingle Beaches", funded by the Ministry of Agri-culture, Fisheries and Food. The departmental nominated. officer for this contract was Mr A J Allison. The company's nominated. project officer was Dr S W Huntington. This report is published on behalf of the Ministry of Agriculture, Fisheries and Food, but the opinions e>rpressed are not necessarily those of the Ministry. @ Crown Copyright 1991 Published by permission of the Controller of Her Majesty's Stationery Office Mathematical model of groSmes on shingle beaches A H Brampton BSc PhD D G Goldberg BA Report SR 276 November 1991 ABSTRACT This report describes the development of a mathematical model of a shingle beach with gro5mes. The development of the beach plan shape is calculated given infornation on its initial position and information on wave conditions just offshore. Different groyne profiles and spacings can be specified, so that alternative gro5me systems can be investigated. Ttre model includes a method for dealing with varying water levels as the result of tidal rise and fall. CONTENTS Page 1. INTRODUCTION I 2. SCOPEOF THE UODEL 3 2.t Model resolution and input conditions 3 2.2 Sediment transport mechanisms 6 2.3 Vertical distribution of sediment transport q 2.4 Wave transformation modelling L0 3. -

By the Sea a Guide to the Coastal Zone of Atlantic
BY THE SEA A GUIDE TO THE COASTAL ZONE OF ATLANTIC CANADA USER’S GUIDE ACKNOWLEDGMENTS: FUNDING: Department of Fisheries and Oceans, COORDINATORS: Roland D. Chiasson, Sabine B. Dietz SUPERVISION & REVISIONS: John A. Legault, Sophie Bastien-Daigle GRAPHICS: Sabine Dietz, Ursula Koch, Elke Leitner, Jean-Raymond Gallien (used with permission from the N.B. Department of Natural Resources and Energy) Également disponible en français. Fisheries Pêches Piper Project/Projet siffleur and Oceans et Océans 4800 Route 11 Tabusintac, N.B. E9H 1J6 Department of Fisheries and Oceans Habitat Management Division 343 Archibald Street Moncton, N.B. E1C 9B6 Prepared by: Corvus Consultants Inc., Tabusintac, N.B., Canada Cat. no. S-23-289/1996 E ©1996 ISBN 0-660-16410-8 THE MODULES MODULE 1 : INTRODUCTORY MODULE MODULE 2 : TO THE HORIZON - THE NEARSHORE MODULE 3 : ESTUARIES MODULE 4 : SALT MARSHES MODULE 5 : TIDAL MUDFLATS MODULE 6 : SANDY BEACHES AND DUNES MODULE 7 : ROCKY SHORES MODULE 8 : COASTAL ISLANDS AND CLIFFS MODULE 9 : COBBLE BEACHES MODULE 10 : COASTAL BOGS MODULE 11 : FRESHWATER BARRIER PONDS MODULE 12 : FJORDS MODULE 13 : ACTIVITIES ABOUT THE MANUAL .......................................................................................................................... 3 1.1. Who is this guide for? 3 1.2. How is this guide organized? 3 1.3. About the ecosystem modules 3 1.4. How to use the ecosystem modules: 4 MODULE DESCRIPTIONS, LEARNING OBJECTIVES, STUDY QUESTIONS AND SUGGESTED ACTIVITIES..................... 5 Activity Suggestions 6 Matrix -

Shoreline Stabilisation
Section 5 SHORELINE STABILISATION 5.1 Overview of Options Options for handling beach erosion along the western segment of Shelley Beach include: • Do Nothing – which implies letting nature take its course; • Beach Nourishment – place or pump sand on the beach to restore a beach; • Wave Dissipating Seawall – construct a wave dissipating seawall in front of or in lieu of the vertical wall so that wave energy is absorbed and complete protection is provided to the boatsheds and bathing boxes behind the wall for a 50 year planning period; • Groyne – construct a groyne, somewhere to the east of Campbells Road to prevent sand from the western part of Shelley Beach being lost to the eastern part of Shelley Beach; • Offshore Breakwater – construct a breakwater parallel to the shoreline and seaward of the existing jetties to dissipate wave energy before it reaches the beach; and • Combinations of the above. 5.2 Do Nothing There is no reason to believe that the erosion process that has occurred over at least the last 50 years, at the western end of Shelley Beach, will diminish. If the water depth over the nearshore bank has deepened, as it appears visually from aerial photographs, the wave heights and erosive forces may in fact increase. Therefore “Do Nothing” implies that erosion will continue, more structures will be threatened and ultimately damaged, and the timber vertical wall become undermined and fail, exposing the structures behind the wall to wave forces. The cliffs behind the wall will be subjected to wave forces and will be undermined if they are not founded on solid rock.