Unit 4: Excretion; Structure of Nephron, Mechanism of Urine Formation, Counter-Current Mechanism
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Role of New Podocyte-Associated Proteins in the Renal Ultrafiltration Barrier
From the DEPARTMENT OF LABORATORY MEDICINE Karolinska Institutet, Stockholm, Sweden ROLE OF NEW PODOCYTE-ASSOCIATED PROTEINS IN THE RENAL ULTRAFILTRATION BARRIER Angelina Schwarz Stockholm 2019 All previously published papers were reproduced with permission from the publisher. Published by Karolinska Institutet. Printed by Universitetsservice US-AB 2019. © Angelina Schwarz, 2019 ISBN 978-91-7831-452-2 Cover: confocal microscopy image of a mouse glomerulus (front) and electron microscopy image of a podocyte (back) Role of new podocyte-associated proteins in the renal ultrafiltration barrier THESIS FOR DOCTORAL DEGREE (Ph.D.) By Angelina Schwarz Principal Supervisor: Opponent: Assoc. Prof. Jaakko Patrakka, MD, PhD Prof. Rachel Lennon, MD, PhD Karolinska Institutet University of Manchester Department of Laboratory Medicine School of Biological Sciences Division of Pathology/ICMC Division of Cell Matrix and Regenerative Medicine Co-supervisor(s): Lwaki Ebarasi, PhD Examination Board: Karolinska Institutet Assoc. Prof. Sergiu-Bogdan Catrina, MD, PhD Department of Laboratory Medicine Karolinska Institutet Division of Pathology/ICMC Department of Molecular Medicine and Surgery Division of Growth and Metabolism Mark Lal, PhD AstraZeneca Prof. Bengt Fellström, MD, PhD Bioscience, Cardiovascular, Renal and Uppsala University Metabolism, Innovative Medicines Biotech Unit Department of Medical Sciences Division of Nephrology Assoc. Prof. Taija Mäkinen, PhD Uppsala University Department of Immunology, Genetics and Pathology Division of Vascular Biology ABSTRACT Chronic kidney disease (CKD) is a major health problem and an economical burden affecting people worldwide. The main causes of CKD are diabetes and hypertension and patient numbers keep increasing. In many cases, CKD is progressive leading to end stage renal disease (ESRD), a condition that can be treated only through chronic dialysis or renal transplantation. -

Development and Structure of the Excretory System the Nephron
Biology 224 Human Anatomy and Physiology II Week 7; Lecture 1; Monday Dr. Stuart S. Sumida Development and Structure of the Excretory System The Nephron DEVELOPMENT AND STRUCTURE OF EXCRETORY SYSTEM EXCRETORY SYSTEM REVIEW • Kidneys derived from INTERMEDIATE MESODERM. • Kidney starts out as a SEGMENTAL STRUCTURE. • Bladder, as part of embryonic gut tube: lining derived from endoderm. • Note, this is EXCRETION, NOT “ELIMINATION.” EARLY KIDNEY DEVELOPMENT • There is a segment of intermediate mesoderm for every body segment. • Earliest kidney appears in the cervical region of the body! (About week 3.) Called the PRONEPHROS. • Develop very close to the gonads. (They battle it out for the nearby ducts.) THE PRONEPHROS • The early anteriorly developed kidney parts. • Has NO EXCRETORY FUNCTION. • Functions to INDUCE DEVELOPMENT of middle segments of intermediate meosderm into the MESONEPHROS. THE MESONEPHROS • Some think it is the functioning embryonic kidney. Some think is has no excretory function. • WE DO KNOW that the duct that attaches to it (THE MESONEPHRIC DUCT) is very important in INDUCING DEVELOPMENT OF THE CAUDAL KIDNEY SEGMENTS (the METANEPHROS). Mesonephric Duct reaches all the way to end of gut tube (cloaca). We need to... • Attach the ducts to the hindend kidney (the metanephros). • Split the bladder away from the gut tube. After the mesonephric duct attaches to cloaca, the embryonic URETER grows from caudal to cranial to attach to mass of metanephric kidney. A septum, the URORECTAL SEPTUM, grows between the more dorsal part of the gut tube and the more ventral part that will become the bladder. Note that attached to the bladder are right and left ueters, the allantois (an extraembryonic membrane). -

The Contribution of Nasal Countercurrent Heat Exchange to Water Balance in the Northern Elephant Seal, Mirounga Angustirostris
J. exp. Biol. 113, 447-454 (1984) 447 Printed in Great Britain © The Company of Biologists Limited 1984 THE CONTRIBUTION OF NASAL COUNTERCURRENT HEAT EXCHANGE TO WATER BALANCE IN THE NORTHERN ELEPHANT SEAL, MIROUNGA ANGUSTIROSTRIS BY ANTHONY C. HUNTLEY, DANIEL P. COSTA Long Marine Laboratory, Center for Marine Studies, University of California, Santa Cruz, U.SA. AND ROBERT D. RUBIN Department of Life Sciences, Santa Rosa Junior College, Santa Rosa, California, U.SA Accepted 29 May 1984 SUMMARY 1. Elephant seals fast completely from food and water for 1-3 months during terrestrial breeding. Temporal countercurrent heat exchange in the nasal passage reduces expired air temperature (Te) below body temperature (Tb)- 2. At a mean ambient temperature of 13'7°C, Te is 20*9 °C. This results in the recovery of 71-5 % of the water added to inspired air. 3. The amount of cooling of the expired air (Tb~Te) and the percentage of water recovery varies inversely with ambient temperature. 4. Total nasal surface area available for heat and water exchange, located in the highly convoluted nasal turbinates, is estimated to be 720 cm2 in weaned pups and 3140 cm2 in an adult male. 5. Nasal temporal countercurrent heat exchange reduces total water loss sufficiently to allow maintenance of water balance using metabolic water production alone. INTRODUCTION Northern elephant seals, Mirounga angustirostis (Gill), are exceptional among pin- nipeds in the duration of their terrestrial breeding fast. During this time, they volun- tarily forgo both food and water while remaining active on the rookery. The length of these fasts varies with age, sex and social status. -

Ward's Renal Lobule Model
Ward’s Renal Lobule Model 470029-444 1. Arcuate artery and vein. 7. Descending thick limb of 12. Collecting tubule. 2. Interlobular artery and vein. Henle's loop. 13. Papillary duct of Bellini. 3. Afferent glomerular arteriole. 8. Thin segment of Henle's 14. Vasa recta. loop. 4. Efferent glomerular arteriole. 15. Capillary bed of cortex (extends 9. Ascending thick limb of through entire cortex). 5. Renal corpuscle (glomerulus Henle's loop. plus Bowman's capsule). 10. Distal convoluted tubule. 16. Capillary bed of medulla (extends 6. Proximal convoluted tubule. 11. Arched connecting tubule. through entire medulla). MANY more banks of glomeruli occur in the cortex than are represented on the model, and the proportionate length of the medullary elements has been greatly reduced. The fundamental physiological unit of the kidney is the nephron, consisting of the glomerulus, Bowman's capsule, the proximal convoluted tubule, Henle's loop, and the distal convoluted tubule. The blood is filtered in the glomerulus, water and soluble substances, except blood proteins, passing into Bowman's capsule in the same proportions as they occur in the blood. In the proximal tubule water and certain useful substances are resorbed from the provisional urine, while some further components may be added to it by secretory activity on the part of the tubular epithelium. In the remainder of the tubule, resorption of certain substances is continued, while the urine is concentrated further by withdrawal of water. The finished urine flows through the collecting tubules without further change. Various kinds of loops occur, varying in length of the thin segment, and in the level to which they descend into the medulla. -

Renal Physiology Background
RENAL PHYSIOLOGY Florida State University Advanced Topics in Biomedical Mathematics MAP5932, Spring 2007 03/16/07 Brinda Pamulapati Goal 1 Background of the kidney 2 Glomerulus 3 Mathematical model of Glomerulus 4 Co-current and counter- current mechanism 5 Mathematical model of the co-current and counter-current mechanism Kidney Kidney and Nephron picture Glomerulus and Bowman's Capsule Mathematical model of the glomerular filter There are 3 pressures that effect the rate of glomerular filtration: 1 the pressure inside the glomerular capillaries that promote filteration(p1) 2 the pressure inside the Bowman's capsule that opposes filtration(p2) 3 the colloidal osmotic pressure of the plasma proteins inside the capillaries that opposes filteration(pi) Schematic diagram of the glomerular filtration(one dimentional) q2 Qi q1 Qe x=0 x=L Mathematical model of the glomerular filter(cont.) dq 1 = K (P " P + ! ) dx f 2 1 c P1 ,P2 = hydrostatic pressure ! c = osmotic pressure of the suspended protei ns and formed elements of the blood K f = capillary filteration rate where osmotic pressure is ! c = RTc Conservation Equation ciQi = cq1 ! c ciQi = q1 ( from osmosis RTc) RT ! c = ! c = RTciQi / q1 Qi ! c = ! i (where ! i = RTci ) q1 Mathematical Model of the Glomerulus dq1 = K (P " P + ! ) ................(20.1) dx f 2 1 c Qi ! c = ! i .......................................................(20.4) qi # Q $ e %! Q & Q ' " e +! ln & i ' =1% K L i f ..............(20.5) Qi & 1%! ' !Qi & ' ( ) Qe =efflux through the efferent arterioles L=length of the filter !=" i /(P1 # P2) Cocurrent and Countercurrent Mechanism What is Cocurrent and Countercurrent Mechanism Why Study about it ? The human kidney use countercurrent exchange to remove water from urine so the body can retain water that was used to move the nitrogenous waste products. -
![L8-Urine Conc. [PDF]](https://docslib.b-cdn.net/cover/4402/l8-urine-conc-pdf-1384402.webp)
L8-Urine Conc. [PDF]
The loop of Henle is referred to as countercurrent multiplier and vasa recta as countercurrent exchange systems in concentrating and diluting urine. Explain what happens to osmolarity of tubular fluid in the various segments of the loop of Henle when concentrated urine is being produced. Explain the factors that determine the ability of loop of Henle to make a concentrated medullary gradient. Differentiate between water diuresis and osmotic diuresis. Appreciate clinical correlates of diabetes mellitus and diabetes insipidus. Fluid intake The total body water Antidiuretic hormone is controled by : Renal excretion of water Hyperosmolar medullary Changes in the osmolarity of tubular fluid : interstitium 1 2 3 Low osmolarity The osmolarity High osmolarity because of active decrease as it goes up because of the transport of Na+ and because of the reabsorbation of water co-transport of K+ and reabsorption of NaCl Cl- 4 5 Low osmolarity because of High osmolarity because of reabsorption of NaCl , also reabsorption of water in reabsorption of water in present of ADH , present of ADH reabsorption of urea Mechanisms responsible for creation of hyperosmolar medulla: Active Co- Facilitated diffusion transport : transport : diffusion : of : Na+ ions out of the Only of small thick portion of the K+ , Cl- and other amounts of water ascending limb of ions out of the thick from the medullary the loop of henle portion of the Of urea from the tubules into the into the medullary ascending limb of inner medullary medullary interstitium the loop of henle collecting -

Thermal Relations 10
CHAPTER Thermal Relations 10 s this bumblebee flies from one flower cluster to another to collect nectar and pollen, temperature matters for the bee in two crucial ways. First, the temperature of the bumblebee’s flight muscles de- A termines how much power they can generate. The flight muscles must be at least as warm as about 35°C to produce enough power to keep the bee airborne; if the muscles are cooler, the bee cannot fly. The second principal way in which temperature matters is that for a bumblebee to maintain its flight muscles at a high enough temperature to fly, the bee must expend food energy to generate heat to warm the muscles. In a warm environment, all the heat required may be produced simply as a by-product of flight. In a cool environment, however, as a bumblebee moves from flower cluster to flower cluster—stopping at each to feed—it must expend energy at an elevated rate even during the intervals when it is not flying, either to keep its flight muscles continually at a high enough temperature to fly or to rewarm the flight muscles to flight temperature if they cool while feeding. Assuming that the flight muscles must be at 35°C for flight, they must be warmed to 10°C above air temperature if the air is at 25°C, but to 30°C above air temperature if the air is at 5°C. Thus, as the air becomes cooler, a bee must expend food energy at a higher and higher rate to generate heat to warm its flight muscles to flight temperature, meaning it must collect food at a higher and higher rate. -

L25 Kidney2 to Post
Vert Phys PCB3743 Kidney 1 Fox Chapter 17 part 1 © T. Houpt, Ph.D. 1 Kidney Function Remove waste chemicals, while reabsorbing nutrients 1. Filter plasma from blood (including water & water-soluble nutrients) 2. Reabsorb Na+ : essential to maintain high extracellular [Na+] 3. Reabsorb H20 : essential to maintain body fluid volume 4. Reabsorb glucose and other nutrients 5. Reabsorb HCO3 / secrete H+ to maintain pH 2 Toxicity of Ammonia (NH3) 1. NH3 -> NH4+ very basic 2. NH3 is metabolic poison • Fish allow NH3 to diffuse into surrounding water • Birds & Reptiles convert NH3 to uric acid, which is not water soluble and is excreted in the feces. • Mammals convert NH3 to urea (CH4N2O), which is non-toxic and water soluble for excretion by kidney 1828: first organic synthesis: the production of urea without living tissue. → AgNCO + NH4Cl (NH2)2CO + AgCl 3 Krebs Cycle http://hyperphysics.phy-astr.gsu.edu/hbase/biology/tca.html 4 Too much ammonia removes a-ketoglutarate from Krebs cycle, so starves cells of ATP http://www.ucl.ac.uk/~ucbcdab/urea/amtox.htm 5 Urea - waste product of excess amino acid metabolism amino acid metabolism toxic! 2 liver ammonia + CO 6 Chapter 17: Anatomy of the Kidney Kidney Function Filter excess and waste chemicals (water soluble) from the blood. (excess water, Na+, urea, glucose > 200 mg/100ml) Kidney Structures cortex (bark): reddish brown, lots of capillaries medulla (inner region): striped with capillaries & collecting ducts; divided into renal pyramids urine -> collecting ducts -> minor calyces -> major calyces calyx; calyces -> renal pelvis-> ureters -> urinary bladder -> urethra “cup” high surface area for exchange, then to storage and outside ren- Latin for kidney nephro- Greek for kidney -uria - problem with urine, e.g. -
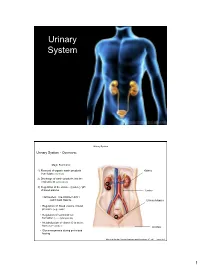
Urinary System
Urinary System Urinary System Urinary System - Overview: Major Functions: 1) Removal of organic waste products Kidney from fluids (excretion) 2) Discharge of waste products into the environment (elimination) 1 3) Regulation of the volume / [solute] / pH 3 of blood plasma Ureter HOWEVER, THE KIDNEY AIN’T JUST FOR PEE’IN… Urinary bladder • Regulation of blood volume / blood pressure (e.g., renin) • Regulation of red blood cell formation (i.e., erythropoietin) 2 • Metabolization of vitamin D to active form (Ca++ uptake) Urethra • Gluconeogenesis during prolonged fasting Marieb & Hoehn (Human Anatomy and Physiology, 8th ed.) – Figure 25.1 1 Urinary System Renal ptosis: Kidneys drop to lower position due Functional Anatomy - Kidney: to loss of perirenal fat Located in the superior lumbar “Bar of soap” region 12 cm x 6 cm x 3 cm 150 g / kidney Layers of Supportive Tissue: Renal fascia: Peritoneal cavity Outer layer of dense fibrous connective tissue; anchors kidney in place Perirenal fat capsule: Fatty mass surrounding kidney; cushions kidney against blows Fibrous capsule: Transparent capsule on kidney; prevents infection of kidney from local tissues Kidneys are located retroperitoneal Marieb & Hoehn (Human Anatomy and Physiology, 8th ed.) – Figure 25.2 Urinary System Functional Anatomy - Kidney: Pyelonephritis: Inflammation of the kidney Pyramids appear striped due to parallel arrangement of capillaries / collecting tubes Renal cortex Renal medulla Renal pyramids Renal papilla Renal columns Renal hilum Renal pelvis • Entrance for blood vessels -

The Distal Convoluted Tubule and Collecting Duct
Chapter 23 *Lecture PowerPoint The Urinary System *See separate FlexArt PowerPoint slides for all figures and tables preinserted into PowerPoint without notes. Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Introduction • Urinary system rids the body of waste products. • The urinary system is closely associated with the reproductive system – Shared embryonic development and adult anatomical relationship – Collectively called the urogenital (UG) system 23-2 Functions of the Urinary System • Expected Learning Outcomes – Name and locate the organs of the urinary system. – List several functions of the kidneys in addition to urine formation. – Name the major nitrogenous wastes and identify their sources. – Define excretion and identify the systems that excrete wastes. 23-3 Functions of the Urinary System Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Diaphragm 11th and 12th ribs Adrenal gland Renal artery Renal vein Kidney Vertebra L2 Aorta Inferior vena cava Ureter Urinary bladder Urethra Figure 23.1a,b (a) Anterior view (b) Posterior view • Urinary system consists of six organs: two kidneys, two ureters, urinary bladder, and urethra 23-4 Functions of the Kidneys • Filters blood plasma, separates waste from useful chemicals, returns useful substances to blood, eliminates wastes • Regulate blood volume and pressure by eliminating or conserving water • Regulate the osmolarity of the body fluids by controlling the relative amounts of water and solutes -

Countercurrent Transport in the Kidney
Ann. Rev. Biophys. Bioeng. 1978. 7:315-39 COUNTERCURRENT +9115 TRANSPORT IN THE KIDNEY 1 John L. Stephenson Section on Theoretical Biophysics, National Heart, Lung and Blood· Institute and Mathematical Research Branch, National Institute of Arthritis, Metabolism, and Digestive Diseases, National Institutes of Health, Bethesda, Maryland 20014 INTRODUCTION Renal physiologists recognized many years ago that the ability of a glomerular kidney to form a concentrated urine was correlated in some way with the possession of a loop of Henle (see 56). It was also recognized that more water than solute must be absorbedfrom the glomerular filtrate to produce a urine more concentrated than plasma. Smith (62, 63) quantitated the relative water reabsorption by noting that if the final urine were to be reduced to isotonicity by the addition or subtraction of water, the total solute excreted by the kidney per minute would occupy a volume of UCM!CpM, where Uis the urine flow, CM is the urine osmolality, and G],M is the plasma osmolality. Since the actual volume occupied by this solute is U. the quantity of water U - UeM! G],M would have to be subtracted when the urine is hypotonic and added when the urine is hypertonic to bring the final urine to isotonicity. Smith called this virtual volume free water for the diluting kidney and negative free water for the concentrating kidney. Since the micropunc ture studies of Richards and his associates had shown that the glomerular filtrate was isotonic and remained so in the proximal tubule (57, 58), it was inferred that in the concentrating kidney the absorption of water in excess of its isotonic complement of solute took place somewhere in the distal nephron. -

Quantitation of Countercurrent Exchange During Passive Absorption from the Dog Small Intestine: Evidence for Marked Species Differences in the Efficiency of Exchange
Quantitation of countercurrent exchange during passive absorption from the dog small intestine: evidence for marked species differences in the efficiency of exchange. J H Bond, … , D G Levitt, M D Levitt J Clin Invest. 1977;59(2):308-318. https://doi.org/10.1172/JCI108642. Research Article The present investigation was designed to quantitatively assess the possible influence of countercurrent exchange on passive absorption from the small intestine of the dog. Villus blood flow was measured with a modification of the microsphere method. Simultaneously, the absorption from the gut lumen of five diffusible gases (H2, He, CH4, 133Xe, and CO) was determined. Villus blood flow averaged 0.247 +/- 0.03 (SEM) ml/min per g. The observed absorption of H2, He, CH4, and 133Xe was only 16.2 +/- 1.8, 12.8 +/- 2.3, 12.0 +/- 1.8, and 15.8 +/- 1.4 %, respectively, of what this villus blood flow could carry away if it reached perfect equilibrium with the luminal gases. This low absorption rate could result from diffusion limitation to absorption or countercurrent exchange. The diffusive permeability of the barrier seperating the luminal gases and villus blood flow was assessed by measuring the absorption rate of CO. Because absorbed CO binds tightly to hemoglobin, it cannot exchange, and when present in low concentrations its uptake is entirely diffusion limited. Knowledge of the diffusion rate through tissue of the unbound gases relative to that of CO made it possible to calculate the degree to which each of the unbound gases should equilibrate with villus tip blood. The percentage equilibration between lumen and blood at the villus tip for H2, He, CH4, […] Find the latest version: https://jci.me/108642/pdf Quantitation of Countercurrent Exchange during Passive Absorption from the Dog Small Intestine EVIDENCE FOR MARKED SPECIES DIFFERENCES IN THE EFFICIENCY OF EXCHANGE JOHN H.