A Single Amino Acid Substitution in the Third Transmembrane Region Has Opposite Impacts on the Selectivity of the Parasiticides
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lindane Lotion USP, 1% RX Only WARNINGS
Lindane Lotion USP, 1% RX Only WARNINGS: Lindane Lotion should only be used in patients who cannot tolerate or have failed first-line treatment with safer medications for the treatment of scabies. (See INDICATIONS AND USAGE.) Neurologic Toxicity Seizures and deaths have been reported following Lindane Lotion use with repeat or prolonged application, but also in rare cases following a single application used according to directions. Lindane lotion should be used with caution for infants, children, the elderly, and individuals with other skin conditions (e.g, atopic dermatitis, psoriasis) and in those who weigh < 110 lbs (50 kg) as they may be at risk of serious neurotoxicity. Contraindications Lindane Lotion is contraindicated in premature infants and individuals with known uncontrolled seizure disorders. Proper Use Instruct patients on the proper use of Lindane Lotion, the amount to apply, how long to leave it on, and avoiding re-treatment. Inform patients that itching occurs after the successful killing of scabies and is not necessarily an indication for re-treatment with Lindane Lotion. (See DOSAGE AND ADMINISTRATION.) DESCRIPTION Lindane Lotion USP, 1%, is an ectoparasiticide and ovicide. In addition to the active ingredient, lindane, it contains glycerol monostearate, cetyl alcohol, stearic acid, trolamine, carrageenan, 2- amino-2-methyl-1-propanol, methylparaben, butylparaben, perfume and water to form a lotion. Lindane is the gamma isomer of 1,2,3,4,5,6-hexachlorocyclohexane having the following structural formula: C6H6Cl6 M.W. 290.83 CLINICAL PHARMACOLOGY Lindane Lotion USP, 1%, is an ectoparasiticide and ovicide effective against Sarcoptes scabiei (scabies). Lindane exerts its parasiticidal action by being directly absorbed into the parasites and 1 their ova. -

Revised Use-Function Classification (2007)
INTERNATIONAL PROGRAMME ON CHEMICAL SAFETY IPCS INTOX Data Management System (INTOX DMS) Revised Use-Function Classification (2007) The Use-Function Classification is used in two places in the INTOX Data Management System: the Communication Record and the Agent/Product Record. The two records are linked: if there is an agent record for a Centre Agent that is the subject of a call, the appropriate Intended Use-Function can be selected automatically in the Communication Record. The Use-Function Classification is used when generating reports, both standard and customized, and for searching the case and agent databases. In particular, INTOX standard reports use the top level headings of the Intended Use-Functions that were selected for Centre Agents in the Communication Record (e.g. if an agent was classified as an Analgesic for Human Use in the Communication Record, it would be logged as a Pharmaceutical for Human Use in the report). The Use-Function classification is very important for ensuring harmonized data collection. In version 4.4 of the software, 5 new additions were made to the top levels of the classification provided with the system for the classification of organisms (items XIV to XVIII). This is a 'convenience' classification to facilitate searching of the Communications database. A taxonomic classification for organisms is provided within the INTOX DMS Agent Explorer. In May/June 2006 INTOX users were surveyed to find out whether they had made any changes to the Use-Function Classification. These changes were then discussed at the 4th and 5th Meetings of INTOX Users. Version 4.5 of the INTOX DMS includes the revised pesticides classification (shown in full below). -

Antibiotics May Eradicate Gastrointestinal Immunodeficiency.8
1122 Letters to the Editor J Neurol Neurosurg Psychiatry: first published as 10.1136/jnnp.54.12.1122 on 1 December 1991. Downloaded from sitivity.? We present a case of OMM without pseudobulbar palsy. The association of vitro.8 Whipple's disease is associated with symptoms of Whipple's disease and with supranuclear gaze paresis with oculomas- immunodeficiency.8 Thus intestinal wall negative peroral jejunoileal biopsies, which ticatory myorhythmia, however, seems to be macrophages are ineffective in phagocytosing indicates the usefulness of laparotomy for pathognomonic of the effects of Whipple's intracellular gram positive bacilli, resulting in jejunoileal biopsies as an alternative to brain disease on the CNS.'2 This led us to perform inability to eliminate chronic infection.9 This biopsy to confirm Whipple's disease. surgical jejunal and mesenteric lymph node suggests that Whipple's disease may be con- A 47 year old woman was admitted in May biopsies rather than a brain biopsy despite sidered as a disease of macrophages.8 The 1988 in a depressed state. In June 1987 she negative endoscopic and numerous peroral periventricular and periaqueductal distribu- had noted progressive visual disturbance. distal jejunal and ileal biopsies. The nor- tion of the CNS involvement in Whipple's Rhythmic elevations of her right upper lip mality ofthe CT and MRI scans also suppor- disease consists of macrophagic infiltration appeared, and later, paroxysmal hypersomnia ted this decision. and subependymal nodules. Such a and considerable weight gain (10 kg). A nine The possibility of brain involvement with- "tumoural" involvement may explain why month course oftreatment for depression was out systemic manifestation in Whipple's dis- antibiotics with good BBB diffusion are not ineffective and her symptoms and signs ease should be kept in mind. -
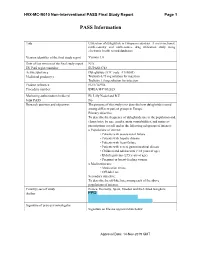
PASS Information
H9X-MC-B010 Non-interventional PASS Final Study Report Page 1 PASS Information Title Utilisation of dulaglutide in European countries: A cross-sectional, multi-country and multi-source drug utilisation study using electronic health record databases Version identifier of the final study report Version 1.0 Date of last version of the final study report N/A EU PAS register number EUPAS13783 Active substance Dulaglutide (ATC code: A10BJ05) Medicinal product(s): Trulicity 0.75 mg solution for injection Trulicity 1.5 mg solution for injection Product reference: EU/1/14/956 Procedure number: EMEA/H/C/002825 Marketing authorisation holder(s) Eli Lilly Nederland B.V. Joint PASS No Research question and objectives The purpose of this study is to describe how dulaglutide is used among different patient groups in Europe. Primary objective: To describe the frequency of dulaglutide use in the population and characterise by age, gender, main comorbidities, and main co- prescriptions overall and in the following subgroups of interest: o Populations of interest: • Patients with severe renal failure • Patients with hepatic disease • Patients with heart failure • Patients with severe gastrointestinal disease • Children and adolescents (<18 years of age) • Elderly patients (≥75 years of age) • Pregnant or breast-feeding women o Medication use: • Medication errors • Off-label use Secondary objective: To describe the off-label use among each of the above populations of interest. Country(-ies) of study France, Germany, Spain, Sweden and the United Kingdom Author PPD Signature of principal investigator Signature on file/see approval date below Approval Date: 14-Nov-2019 GMT H9X-MC-B010 Non-interventional PASS Final Study Report Page 2 Marketing Authorisation Holder Marketing authorisation holder (MAH) Eli Lilly Nederland B.V. -

A Saprolegnia Parasitica Challenge System for Rainbow Trout: Assessment of Pyceze As an Anti-Fungal Agent for Both Fish and Ova
DISEASES OF AQUATIC ORGANISMS Published May 12 Dis Aquat Org l A Saprolegnia parasitica challenge system for rainbow trout: assessment of Pyceze as an anti-fungal agent for both fish and ova T. G.Pottinger*, J. G. Day NERC Institute of Freshwater Ecology, Windermere Laboratory, Far Sawrey, Ambleside, Cumbria, LA22 OLP, United Kingdom ABSTRACT: A reproducible Saprolegnia parasitica spore delivery system was developed and demon- strated to be effective in providing a sustained spore challenge for up to 10 d. Treatment of rainbow trout with slow-release intraperitoneal implants containing cortisol resulted in chronically elevated blood cortisol levels and rendered the fish susceptible to infection by S. parasitica when exposed to the spore challenge. Sham-implanted fish were not susceptible to infect~on.Bronopol (2-bromo-2-nitro- propane-1,3-dol), formulated as Pyceze, was effective in protecting predisposed fish from infection by S. parasitica when administered as a daily bathlflush treatment at concentrations of 15 mg I-' and greater. Pyceze was also demonstrated to protect fertilised rainbow trout ova from S. parasitica chal- lenge when administered as a daily bath/flush treatment at concentrations of between 30 and 100 mg 1-l. Pyceze appears to qualify as a safe and effective replacement for malachite green and formalin in the prevention of fungal infections in the aquaculture environment. KEY WORDS: Saprolegnia . Fungal infection . Salmonid . Bronopol - Pyceze . Cortisol INTRODUCTION chite green in combating mycotic infections of fish and fish eggs, but is safer for the operator, the fish, and Mycotic infections of farmed fish, primarily by water the environment. Among the alternative compounds moulds or pseudofungi of the genus Saprolegnia, rep- tested for antifungal activity are sodium chloride resent a significant economic and welfare problem. -

Study Protocol
STUDY PROTOCOL FOR COMPASSIONATE AQUACULTURE INVESTIGATIONAL NEW ANIMAL DRUG (INAD) EXEMPTION FOR 35% PEROX-AID® (HYDROGEN PEROXIDE) FOR THE CONTROL OF ECTOPARASITES (INAD #11-669) Sponsor: U.S. Fish and Wildlife Service, Division of the National Fish Hatchery System ______________________ ___________________ Sponsor Signature Date Approved Manufacturer: Eka Chemicals Inc. 1775 West Oak Commons Court Marietta, Georgia 30062-2254 Facility for Coordination of 35% PEROX AID® INAD: Aquatic Animal Drug Approval Partnership Program 4050 Bridger Canyon Road Bozeman, Mt 59715 Proposed Starting Date: December 1, 2007 Proposed Ending Date: November 30, 2012 Study Director: Mr. Jim Bowker _________________________ ________________ Study Director Signature Date Clinical Field Trial Location and Trial Number: Facility ______________________________________________________________ Type or Print Name Investigator____________________________________________________________ Type or Print Name __________________________________________ ________________________ Investigator Signature Date 1 I. STUDY ID AND TITLE 3 II. SPONSOR 3 III. INVESTIGATORS/FACILITIES 4 IV. PROPOSED STARTING AND COMPLETION DATES: 4 V. BACKGROUND/PURPOSE 4 VI. SPECIFIC OBJECTIVES 6 VII. MATERIALS 7 VIII. EXPERIMENTAL UNIT 10 IX. ENTRANCE CRITERIA 10 X. TREATMENT GROUPS 11 XI. TREATMENT SCHEDULES 12 XII. TREATMENT RESPONSE PARAMETERS 15 XIII. FORMS FOR DATA COLLECTION 15 XIV. RECORD KEEPING PROCEDURES 16 XV. DISPOSITION OF INVESTIGATIONAL ANIMALS 16 XVI. DISPOSITION OF INVESTIGATIONAL -

Laboratory and Field Studies to Investigate the Efficacy of a Novel
Kryda et al. Parasites Vectors (2019) 12:445 https://doi.org/10.1186/s13071-019-3702-6 Parasites & Vectors RESEARCH Open Access Laboratory and feld studies to investigate the efcacy of a novel, orally administered combination product containing moxidectin, sarolaner and pyrantel for the prevention of heartworm disease (Diroflaria immitis) in dogs Kristina Kryda1*, Robert H. Six1, Kelly F. Walsh1, Susan J. Holzmer1, Sara Chapin1, Sean P. Mahabir1, Melanie Myers1, Tammy Inskeep1, Jady Rugg1, Blair Cundif1, Aleah Pullins1, Michael Ulrich2, John W. McCall3, Tom L. McTier1 and Steven J. Maeder1 Abstract Background: Diroflaria immitis is a flarial parasite of dogs that can cause serious or fatal cardiopulmonary disease. Three studies were conducted to evaluate the efcacy and safety of monthly treatment with moxidectin in a chew- able tablet product in combination with sarolaner and pyrantel to prevent heartworm disease in dogs after experi- mental challenge and in a clinical feld study in the USA. Methods: In two laboratory studies, dogs (8 per group) that had been inoculated 30 days prior with 50 third-stage D. immitis larvae were randomized to treatment on Day 0 with placebo or combination product, at the minimum dose of 24 µg/kg moxidectin, 2 mg/kg sarolaner and 5 mg/kg pyrantel (as pamoate salt). Study 2 also included groups treated with tablets containing moxidectin-alone (24 µg/kg) or sarolaner-alone (2 mg/kg). Efcacy was evaluated ~ 5 months after inoculation by adult heartworm counts at necropsy. In the feld study, 410 dogs 8 weeks-old from 23 USA veterinary clinics were treated for 11 months with either combination product at 24–48 µg/kg≥ moxidectin, 2–4 mg/kg sarolaner and 5–10 mg/kg pyrantel (n 272) or Heartgard® Plus (ivermectin/pyrantel) at the label recom- mended dose rate (n 138). -

Pharmaceuticals As Environmental Contaminants
PharmaceuticalsPharmaceuticals asas EnvironmentalEnvironmental Contaminants:Contaminants: anan OverviewOverview ofof thethe ScienceScience Christian G. Daughton, Ph.D. Chief, Environmental Chemistry Branch Environmental Sciences Division National Exposure Research Laboratory Office of Research and Development Environmental Protection Agency Las Vegas, Nevada 89119 [email protected] Office of Research and Development National Exposure Research Laboratory, Environmental Sciences Division, Las Vegas, Nevada Why and how do drugs contaminate the environment? What might it all mean? How do we prevent it? Office of Research and Development National Exposure Research Laboratory, Environmental Sciences Division, Las Vegas, Nevada This talk presents only a cursory overview of some of the many science issues surrounding the topic of pharmaceuticals as environmental contaminants Office of Research and Development National Exposure Research Laboratory, Environmental Sciences Division, Las Vegas, Nevada A Clarification We sometimes loosely (but incorrectly) refer to drugs, medicines, medications, or pharmaceuticals as being the substances that contaminant the environment. The actual environmental contaminants, however, are the active pharmaceutical ingredients – APIs. These terms are all often used interchangeably Office of Research and Development National Exposure Research Laboratory, Environmental Sciences Division, Las Vegas, Nevada Office of Research and Development Available: http://www.epa.gov/nerlesd1/chemistry/pharma/image/drawing.pdfNational -

United States Patent 19 11 Patent Number: 5,446,070 Mantelle (45) Date of Patent: "Aug
USOO544607OA United States Patent 19 11 Patent Number: 5,446,070 Mantelle (45) Date of Patent: "Aug. 29, 1995 54 COMPOST ONS AND METHODS FOR 4,659,714 4/1987 Watt-Smith ......................... 514/260 TOPCAL ADMNSTRATION OF 4,675,009 6/1987 Hymes .......... ... 604/304 PHARMACEUTICALLY ACTIVE AGENTS 4,695,465 9/1987 Kigasawa .............................. 424/19 4,748,022 5/1988 Busciglio. ... 424/195 75 Inventor: Juan A. Mantelle, Miami, Fla. 4,765,983 8/1988 Takayanagi. ... 424/434 4,789,667 12/1988 Makino ............ ... 514/16 73) Assignee: Nover Pharmaceuticals, Inc., Miami, 4,867,970 9/1989 Newsham et al. ... 424/435 Fla. 4,888,354 12/1989 Chang .............. ... 514/424 4,894,232 1/1990 Reul ............. ... 424/439 * Notice: The portion of the term of this patent 4,900,552 2/1990 Sanvordeker .... ... 424/422 subsequent to Aug. 10, 2010 has been 4,900,554 2/1990 Yanagibashi. ... 424/448 disclaimed. 4,937,078 6/1990 Mezei........... ... 424/450 Appl. No.: 112,330 4,940,587 7/1990 Jenkins ..... ... 424/480 21 4,981,875 l/1991 Leusner ... ... 514/774 22 Filed: Aug. 27, 1993 5,023,082 6/1991 Friedman . ... 424/426 5,234,957 8/1993 Mantelle ........................... 514/772.6 Related U.S. Application Data FOREIGN PATENT DOCUMENTS 63 Continuation-in-part of PCT/US92/01730, Feb. 27, 0002425 6/1979 European Pat. Off. 1992, which is a continuation-in-part of Ser. No. 0139127 5/1985 European Pat. Off. 813,196, Dec. 23, 1991, Pat. No. 5,234,957, which is a 0159168 10/1985 European Pat. -

Reflection Paper on Resistance in Ectoparasites
1 13 September 2018 2 EMA/CVMP/EWP/310225/2014 3 Committee for Medicinal Products for Veterinary Use (CVMP) 4 Reflection paper on resistance in ectoparasites 5 Draft Draft agreed by Efficacy Working Party (EWP-V) May 2018 Adopted by CVMP for release for consultation September 2018 Start of public consultation 21 September 2018 End of consultation (deadline for comments) 31 August 2019 6 Comments should be provided using this template. The completed comments form should be sent to [email protected] 7 Keywords Ectoparasites, resistance to ectoparasiticides 30 Churchill Place ● Canary Wharf ● London E14 5EU ● United Kingdom Telephone +44 (0)20 3660 6000 Facsimile +44 (0)20 3660 5555 Send a question via our website www.ema.europa.eu/contact An agency of the European Union © European Medicines Agency, 2018. Reproduction is authorised provided the source is acknowledged. 8 Reflection paper on resistance in ectoparasites 9 Table of contents 10 1. Introduction ....................................................................................................................... 4 11 2. Definition of resistance ...................................................................................................... 4 12 3. Current state of ectoparasite resistance ............................................................................ 4 13 3.1. Ticks .............................................................................................................................. 4 14 3.2. Mites ............................................................................................................................. -
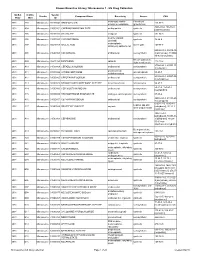
Known Bioactive Library: Microsource 1 - US Drug Collection
Known Bioactive Library: Microsource 1 - US Drug Collection ICCB-L ICCB-L Vendor Vendor Compound Name Bioactivity Source CAS Plate Well ID antifungal, inhibits Penicillium 2091 A03 Microsource 00200046 GRISEOFULVIN 126-07-8 mitosis in metaphase griseofulvum 3505-38-2, 486-16-8 2091 A04 Microsource 01500161 CARBINOXAMINE MALEATE antihistaminic synthetic [carbinoxamine] 2091 A05 Microsource 00200331 SALSALATE analgesic synthetic 552-94-3 muscle relaxant 2091 A06 Microsource 01500162 CARISOPRODOL synthetic 78-44-4 (skeletal) antineoplastic, 2091 A07 Microsource 00210369 GALLIC ACID insect galls 149-91-7 astringent, antibacterial 66592-87-8, 50370-12- 2091 A08 Microsource 01500163 CEFADROXIL antibacterial semisynthetic 2 [anhydrous], 119922- 89-9 [hemihydrate] Rheum palmatum, 2091 A09 Microsource 00211468 DANTHRON cathartic 117-10-2 Xyris semifuscata 27164-46-1, 25953-19- 2091 A10 Microsource 01500164 CEFAZOLIN SODIUM antibacterial semisynthetic 9 [cefazolin] glucocorticoid, 2091 A11 Microsource 00300024 HYDROCORTISONE adrenal glands 50-23-7 antiinflammatory 64485-93-4, 63527-52- 2091 A12 Microsource 01500165 CEFOTAXIME SODIUM antibacterial semisynthetic 6 [cefotaxime] 2091 A13 Microsource 00300029 DESOXYCORTICOSTERONE ACETATE mineralocorticoid adrenocortex 56-47-3 58-71-9, 153-61-7 2091 A14 Microsource 01500166 CEPHALOTHIN SODIUM antibacterial semisynthetic [cephalothin] 2091 A15 Microsource 00300034 TESTOSTERONE PROPIONATE androgen, antineoplastic semisynthetic 57-85-2 24356-60-3, 21593-23- 2091 A16 Microsource 01500167 CEPHAPIRIN SODIUM -

Assessment of the Anti-Feeding and Insecticidal
Assessment of the anti-feeding and insecticidal effects of the combination of dinotefuran, permethrin and pyriproxyfen (Vectra((R)) 3D) against Triatoma infestans on rats D. Tahir, B. Davoust, M. Varloud, J. -M. Berenger, Didier Raoult, L. Almeras, Philippe Parola To cite this version: D. Tahir, B. Davoust, M. Varloud, J. -M. Berenger, Didier Raoult, et al.. Assessment of the anti-feeding and insecticidal effects of the combination of dinotefuran, permethrin and pyriproxyfen (Vectra((R)) 3D) against Triatoma infestans on rats. Medical and Veterinary Entomology, Wiley, 2017, 31 (2), pp.132-139. 10.1111/mve.12206. hal-01573738 HAL Id: hal-01573738 https://hal.archives-ouvertes.fr/hal-01573738 Submitted on 24 Apr 2018 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Medical and Veterinary Entomology (2017) 31, 132–139 doi: 10.1111/mve.12206 Assessment of the anti-feeding and insecticidal effects of the combination of dinotefuran, permethrin and pyriproxyfen (Vectra® 3D) against Triatoma infestans on rats D. TAHIR1, B. DAVOUST1, M. VARLOUD2, J.-M. BERENGER1, D. RAOULT1, L. ALMERAS1 andP.PAROLA1 1Unité de Recherche en Maladies Infectieuses et Tropicales Emergentes (URMITE), UM63, Centre National de la Recherche Scientifique (CNRS) 7278, Institut de Recherche pour le Développement (IRD) 198 (Dakar), Institut National de la Santéetdela Recherche Médicale (INSERM) 1095, Aix-Marseille University, Marseille, France and 2Ceva Santé Animale SA, Libourne, France Abstract.