Development and Application of Passive Integrated
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Flood Frequency Analyses for New Brunswick Rivers Canadian Technical Report of Fisheries and Aquatic Sciences 2920
Flood Frequency Analyses for New Brunswick Rivers Aucoin, F., D. Caissie, N. El-Jabi and N. Turkkan Department of Fisheries and Oceans Gulf Region Oceans and Science Branch Diadromous Fish Section P.O. Box 5030, Moncton, NB, E1C 9B6 2011 Canadian Technical Report of Fisheries and Aquatic Sciences 2920 Canadian Technical Report of Fisheries and Aquatic Sciences Technical reports contain scientific and technical information that contributes to existing knowledge but which is not normally appropriate for primary literature. Technical reports are directed primarily toward a worldwide audience and have an international distribution. No restriction is placed on subject matter and the series reflects the broad interests and policies of Fisheries and Oceans, namely, fisheries and aquatic sciences. Technical reports may be cited as full publications. The correct citation appears above the abstract of each report. Each report is abstracted in the data base Aquatic Sciences and Fisheries Abstracts. Technical reports are produced regionally but are numbered nationally. Requests for individual reports will be filled by the issuing establishment listed on the front cover and title page. Numbers 1-456 in this series were issued as Technical Reports of the Fisheries Research Board of Canada. Numbers 457-714 were issued as Department of the Environment, Fisheries and Marine Service, Research and Development Directorate Technical Reports. Numbers 715-924 were issued as Department of Fisheries and Environment, Fisheries and Marine Service Technical Reports. The current series name was changed with report number 925. Rapport technique canadien des sciences halieutiques et aquatiques Les rapports techniques contiennent des renseignements scientifiques et techniques qui constituent une contribution aux connaissances actuelles, mais qui ne sont pas normalement appropriés pour la publication dans un journal scientifique. -

Septembre 2016 Utilisation Du Guide Des Points
MARCHANDISES GÉNÉRALES Points de Service Day & Ross Marchandises Générales Septembre 2016 Utilisation du guide des points Les points de parité et le guide des points publiés Tout tarif appliqué à un point associé à un point de parité ou basé sur celui-ci sera le même que le tarif du point de base. Par exemple, Dieppe, au Nouveau-Brunswick, PROV VILLE TERMINAL NOTE 1 NOTE 2 POINT DE BASE dont le terminal de livraison est Moncton (MTN), est associé à Moncton, au Nouveau-Brunswick. NB DIEPPE MTN MONCTON Par conséquent, tous les taux et frais de la grille tarifaire s’appliquant à Moncton, s’appliqueront également à Dieppe. Changements et exceptions Cette application ne peut pas être utilisée lorsque le tarif est publié pour un point autre qu’un point de parité tel que Dieppe. Par conséquent, le point précisé dans le tarif du client aura préséance, que Moncton soit également publié dans ce tarif ou non. Les tarifs des points ultérieurs de Corner Brook à Labrador sont saisonniers. Les tarifs aériens s’appliqueront en hiver si le service est oert, en absence du service de traversier, sinon les expéditions à destination de Labrador ne seront pas acceptées, à moins que le service puisse être assuré par le terminal de Québec. Les tarifs vers certains points du nord-ouest, de Nunavut, du Yukon et du Labrador peuvent changer en hiver et pendant la période de débâcle du printemps à cause des fermetures de route et de l’interruption du service de traversier. Le service aérien sera oert sur demande. Notes La note 1 du guide des points porte sur les jours de service supplémentaires qui pourraient s’avérer nécessaires. -

Atlantic Salmon Southwest New Brunswick Outer Fundy SFA 23
Fisheries Pêches and Oceans et Océans DFO Science Maritimes Region Stock Status Report D3-13 Atlantic Salmon Southwest New Brunswick outer Fundy SFA 23 Background Atlantic salmon inhabit 7 major rivers of the “outer Fundy” area of southwest New Brunswick. The rivers are bounded by and include the Saint John and St. Croix. Salmon stocks of these rivers are characterized by a maiden MSW (multi-sea- winter; large) component that, unlike inner Fundy stocks, migrates to the Labrador Sea and a 1SW (one-sea-winter; small) component that is known to venture to the south and east coasts of insular Newfoundland. This report summarizes the status of salmon stocks of the Saint John River above The Fishery Mactaquac, the Nashwaak, Kennebecasis and Hammond tributaries below Mactaquac and the Magaguadavic and St. Croix rivers. Within southwest New Brunswick, Aborig- inal peoples, including five First Nations, Stock characteristics vary between outer Fundy rivers, e.g., wild MSW fish comprise about 40% of total wild returns to were allocated 3,795 1SW fish, mostly from above Mactaquac, the Nashwaak and the Magaguadavic stocks returning to Mactaquac. Estimated rivers and 70% of wild returns to the St. Croix River. MSW salmon are 75- 85% female on the Saint John and 60% female harvests were 675 1SW and 285 MSW fish. on the St. Croix and provide most of the egg deposition. Wild In 1995, a fall harvest after a summer-long 1SW salmon are 15% female above Mactaquac, 40% female in the Nashwaak and 50% female in the St. Croix. About 20% closure was estimated at only 75 fish. -

Status of Atlantic Salmon Stocks of Southwest New Brunswick, 1996
Department of Fisheries and Oceans . Ministère des pêches et océan s Canadian Stock Assessment Secretariat Secrétariat canadien pour l'évaluation des stocks Research Document 97/27 Document de recherche 97/2 7 Not to be cited without Ne pas citer sans permission of the authors ' autorisation des auteurs ' Status of Atlantic salmon stocks of southwest New Brunswick, 199 6 by T.L. Marshall ' Science Branch, Maritimes Region Dept. Fisheries and Oceans P .O. Box 550, Halifax, N.S., B3J 2S7/ and R.A. Jones Science Branch, Maritimes Region Dept. Fisheries and Oceans P.O. Box 5030, Moncton, N.B., E1 C 9B6 and T. Pettigrew N.B. Dept . Natural Resources and Energy P.O. Box 150. Hampton, N.B., EOG 1ZO 1 This series documents the scientific basis for ' La présente série documente les bases the evaluation of fisheries resources in scientifiques des évaluations des ressources Canada. As such, it addresses the issues of halieutiques du Canada. Elle traite des the day in the time frames required and the problèmes courants selon les échéanciers documents it contains are not intended as dictés. Les documents qu'elle contient ne definitive statements on the subjects doivent pas être considérés comme des .addressed but rather as progress reports on énoncés définitifs sur les sujets traités, mais ongoing investigations . plutôt comme des rapports d'étape sur les études en cours . Research documents are produced in the Les documents de recherche sont publiés dans official language in which they are provided to la langue officielle utilisée dans le manuscrit the Secretariat. envoyé au secrétariat . 2 TABLE OF CONTENTS Abstract . -

GST/HST Incremental Federal Rebate For
GST/HST Incremental Federal Rebate for Municipalities Report - January 1, 2013 to December 31 2013 Rapport sur les remboursements fédéraux additionnels de la TPS/TVH aux municipalités - Du 1er janvier au 31 décembre 2013 POSTAL CODE/ PAYMENT AMOUNT/ LEGAL NAME/DÉNOMINATION SOCIALE CITY/VILLE CODE POSTAL MONTANT DU PAIEMENT Alberta/Alberta 1016819 ALBERTA LTD. ARDROSSAN T8E $274.42 1203213 ALBERTA LTD. DEVON T9G $9,957.14 1207175 ALBERTA LTD. BEZANSON T0H $6,961.79 124 STREET AND AREA BUSINESS ASSOCIATION EDMONTON T5M $2,006.39 340715 ALBERTA SOCIETY GRANDE PRAIRIE T8V $5,222.56 4D HOLDINGS LTD. WORSLEY T0H $1,542.39 676853 ALBERTA LTD RED EARTH CREEK T0G $1,653.36 952133 ALBERTA LTD. LETHBRIDGE T1J $2,743.32 967204 ALBERTA LTD. ARDROSSAN T8E $175.01 ABBY ROAD HOUSING CO-OPERATIVE LTD EDMONTON T6G $4,741.03 AC PUREWATER HAULING SERVICES LTD SPRUCE GROVE T7X $1,210.26 ACADIA FOUNDATION OYEN T0J $70,104.98 ACCM BENEVOLENT ASSOCIATION EDMONTON T6C $8,936.69 ACTON HOUSE HOUSING CO-OPERATIVE LIMITED ROCKY MOUNTAIN HOUSE T4T $635.67 AIRDRIE HOUSING LIMITED AIRDRIE T4B $917.47 ALBERTA AVENUE BUSINESS REVITALIZATION ZONE ASSOCIATION EDMONTON T5B $1,270.03 ALBERTA CAPITAL REGION WASTEWATER COMMISSION FORT SASKATCHEWAN T8L $441,060.12 ALBERTA SOUTHWEST REGIONAL ALLIANCE LTD. PINCHER CREEK T0K $5,174.38 ALBERTA WATER AUTHORITY CORP. RED DEER T4P $3,325.43 ALTERNATE CHOICE WATER HAULING LTD. EDMONTON T5W $7,825.63 ANDRE'S WATER DELIVERY LTD LAC LA BICHE T0A $6,719.98 ANSGAR LUTHERAN HOUSING EDMONTON T5K $13,275.34 AQUALINE EXPRESS (2011) LTD. -

Appendix 9-1 Municipal and Regional Authorities and Emergency
CA PDF Page 1 of 35 Energy East Pipeline Ltd. Energy East Project Consolidated Application Volume 9: Community Engagement Appendix 9-1 Municipal and Regional Authorities and Emergency Response Stakeholders May 2016 CA PDF Page 2 of 35 Appendix 9-1 Energy East Pipeline Ltd. Engaged Municipalities and Emergency Consolidated Application Response Stakeholders Table 9-1: Engaged Municipalities and Regional Authorities Engaged Municipalities and Regional Authorities – Alberta Alberta Association of Municipal Districts & Counties (AAMDC) Alberta Rural Municipalities Administrators Association (ARMAA) Alberta Urban Municipalities Association (AUMA) City of Camrose City of Medicine Hat County of Newell County of Paintearth County of Stettler Cypress County Flagstaff County MD of Acadia MD of Provost MD of Wainwright Special Areas Board Town of Castor Town of Coronation Town of Daysland Town of Hanna Town of Hardisty Town of Killam Town of Oyen Town of Provost Town of Sedgewick Town of Vermilion Town of Viking Town of Wainwright Village of Alliance Village of Amisk Village of Cereal Village of Consort Village of Czar Village of Empress Village of Forestburg Village of Galahad Village of Halkirk Village of Heisler Village of Hughenden Village of Lougheed Village of Strome May 2016 Page 1 of 34 CA PDF Page 3 of 35 Appendix 9-1 Engaged Municipalities and Emergency Energy East Pipeline Ltd. Response Stakeholders Consolidated Application Table 9-1: Engaged Municipalities and Regional Authorities (cont’d) Engaged Municipalities and Regional Authorities -

Atlantic Salmon SFA 23
Fisheries Pêches and Oceans et Océans DFO Science Maritimes Region Stock Status Report D3-13(1998) Atlantic Salmon Southwest New Brunswick outer-Fundy SFA 23 Background Atlantic salmon inhabit 7 major rivers of the “outer- Fundy” area of southwest New Brunswick. The rivers are bounded by and include the Saint John and St. Croix. Salmon of these rivers are characterized by a maiden MSW (multi-sea-winter; large) component that, unlike inner-Fundy stocks, migrates to the Labrador Sea and a 1SW (one-sea- winter; small) component that is known to venture to the south and east coasts of insular Newfoundland. Releases in 1997 numbered about 310,000 This report summarizes the status of salmon stocks smolts, 700,000 fry and 422,000 age 0+ fall of the Saint John River above Mactaquac, the parr. Nashwaak, Kennebecasis and Hammond rivers below Mact-aquac and the Magaguadavic and St. The New Brunswick Atlantic salmon Croix rivers. aquaculture industry is concentrated in the Passamaquoddy Bay (Fundy Isles) area. Stock characteristics vary between outer-Fundy Growers produced an estimated 18,500t of rivers, e.g., wild MSW fish comprise about 40% of Atlantic salmon in 1997. An outbreak of ISA total wild returns to above Mactaquac, the (Infectious Salmon Anemia virus), a causative Nashwaak and the Magaguadavic rivers and 70% of agent of HKS (Hemorrhagic Kidney Syndrome) wild returns to the St. Croix River. MSW salmon are mostly affected fish that were to have been 75-95% female on the Saint John and 60% female on harvested in 1998. Escapes in 1997 were not the St. -
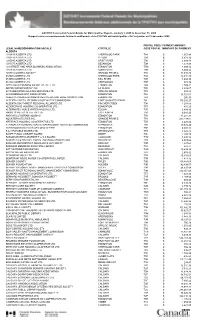
Code Postal Payment Amount
GST/HST Incremental Federal Rebate for Municipalities Report - January 1, 2009 to December 31, 2009 Rapport sur les remboursements fédéraux additionnels de la TPS/TVH aux municipalités - Du 1er janvier au 31 décembre 2009 POSTAL CODE/ PAYMENT AMOUNT/ LEGAL NAME/DÉNOMINATION SOCIALE CITY/VILLE CODE POSTAL MONTANT DU PAIEMENT ALBERTA 1016819 ALBERTA LTD. SHERWOOD PARK T8A $ 1,557.48 1203213 ALBERTA LTD. DEVON T9G $ 5,474.16 1205585 ALBERTA LTD. SPIRIT RIVER T0H $ 3,089.48 1207175 ALBERTA LTD. BEZANSON T0H $ 1,573.46 124 STREET AND AREA BUSINESS ASSOCIATION EDMONTON T5M $ 1,599.12 1287616 ALBERTA LTD. BONNYVILLE T9N $ 5,304.92 340715 ALBERTA SOCIETY GRANDE PRAIRIE T8V $ 41,540.76 882020 ALBERTA LTD. SHERWOOD PARK T8A $ 14,891.39 919055 ALBERTA LTD KEG RIVER T0H $ 3,694.90 967204 ALBERTA LTD. ARDROSSAN T8E $ 873.79 ABBY ROAD HOUSING CO-OPERATIVE LTD EDMONTON T6G $ 1,799.41 ABYSS WATER SUPPLY INC. LA GLACE T0H $ 3,439.07 AC PUREWATER HAULING SERVICES LTD SPRUCE GROVE T7X $ 989.83 ACCM BENEVOLENT ASSOCIATION EDMONTON T6C $ 13,723.03 ALBERTA AVENUE BUSINESS REVITALIZATION ZONE ASSOCIATION EDMONTON T5G $ 296.95 ALBERTA CAPITAL REGION WASTEWATER COMMISSION FORT SASKATCHEWAN T8L $ 175,030.27 ALBERTA SOUTHWEST REGIONAL ALLIANCE LTD. PINCHER CREEK T0K $ 7,293.89 ALDERGROVE HOUSING CO-OPERATIVE LTD EDMONTON T5T $ 825.95 ALTERNATE CHOICE WATER HAULING LTD. EDSON T7E $ 6,894.10 ANDRE'S WATER DELIVERY LTD LAC LA BICHE T0A $ 9,010.95 ANSGAR LUTHERAN HOUSING EDMONTON T5K $ 14,281.38 AQUATERA UTILITIES INC. GRANDE PRAIRIE T8V $ 403,739.07 ARTSPACE HOUSING CO-OPERATIVE LTD EDMONTON T5H $ 4,220.04 ATHABASCA REGIONAL WASTE MANAGEMENT SERVICES COMMISSION ATHABASCA T9S $ 19,615.45 AVENWOOD/BUCHANAN SENIORS CENTRE EDMONTON T5B $ 5,898.18 B & V POTABLE WATER LTD. -

Annual Report 2019 Canada Day Arts & Culture Park 2019 Content
Annual Report 2019 Canada Day Arts & Culture Park 2019 Content 2 Message from the Mayor 3-4 Your Council/Remuneration 5 Community Profile 6 Regional Contributions 7 Civic Relations 8 Town Revenues 9-10 Municipal Services 11 Water & Wastewater Utility 12-14 2019 Highlights 15 Appendixes 22 2019 Financial Statements This annual report is prepared pursuant to Section 105 of the Local Governance Act, and Regulation 2018-54. This report contains general information about Quispamsis such as its tax base, tax rate and user charges, as well as more detailed information regarding town council, the provision of grants, and types and cost of services provided. The 2019 Audited Financial Statements are appended as Appendix V. Message from the Mayor Mayor Gary Clark Quispamsis is the sixth-largest municipality in New Brunswick. Did you know our population is greater than half of the province’s eight cities? For the second year in a row, Quispamsis and Rothesay, the towns protected by the Kennebecasis Regional Police Force, have been deemed to be the safest place to live in Canada, an honour we share with Petawawa, ON. That designation is based on crime severity index numbers compiled by Statistics Canada. Quispamsis residents are resilient. Once again this year, we dealt with extreme flooding conditions, and we came together to help our friends and neighbours. I believe we have grown stronger thanks to this outpouring of community support. I encourage everyone to follow us on social media - @TownofQuispamsis on Facebook and @Quispamsis on Twitter - to keep up-to-date on everything that is happening in our town. -

Hampton Historic Homes Walking Tour
STAGECOACH HOUSE GREEN BRIDGE 10 1076 Main Street 11 Scenic pedestrian river passage This house is possibly the oldest in Hampton. It is believed To complete your walking tour, get the best view of the that it was built by the Loyalist, Abner Hampton, in the early Kennebecasis River with a short stroll across the pedestrian days of settlement. walkway on the ‘Green Bridge’ (Kennebecasis River Bridge HISTORIC HOMES Over the years the #2). This is a spectacular spot for photos with views of the building has served river, birds, and wildlife. Built in 1932, the bridge has had four many purposes; predecessors. The first bridge (wooden) was washed out by WALKING TOUR stagecoach stop, flooding in 1854. Two other wooden bridges were built be- Just off the beaten path is a small town carriage shop, meat tween 1854 and 1891. Then the first steel bridge was erect- with big stories. market, bicycle re- ed. Prior to 1854, a ferry was used to transport people and pair shop, ice cream agricultural produce from farms along the opposite shore. A place of outdoor adventure parlour, funeral Today when you cross the bridge, a short two minute walk and at-home hospitality home, and now a family residence. will bring you to Kredl’s Corner Market. This local favourite HAMPTON, NEW BRUNSWICK After WWII, the pasture adjacent to the Stage Coach House is open year round A place of hometown heroes was used to house the mascot of the 8th Hussars Regiment, with farm fresh and homegrown goodness. a horse named Princess Louise. -

Renewal Application of Kelly Cove Salmon Ltd. for AQ#1205 1. Overview
Findings and Decision- Renewal Application of Kelly cove Salmon Ltd. for AQ#1205 1. Overview: On October 8, 2019, the Nova Scotia Department of Fisheries and Aquaculture (NSDFA) received an application from Kelly Cove Salmon Ltd. to renew Aquaculture Licence and Lease #1205 (AQ#1205), as described below: Table 1. Description of Aquaculture Licence and Lease #1205 Type: Marine Size: 3.99HA Number: AQ#1205 Cultivation Method: Marine Cage Applicant: Kelly Cove Salmon Ltd. Species: Atlantic salmon and Rainbow trout Location: Liverpool Bay, Queens County Proposed Term: 10 year Licence/20 year Lease 2. History AQ#1205 was issued to Sun Salmon Ltd. on March 27, 2000 for a ten-year term (April 1, 2000 to April 1, 2010). AQ#1205 was assigned to Ocean Trout Farms Inc. on July 16, 2004. AQ#1205 was renewed on July 14, 2010 for a five-year term (April 2, 2010 to April 1, 2015). AQ#1205 was assigned to Kelly Cove Salmon Ltd. on May 17, 2012. AQ#1205 was renewed on July 14, 2015 for a five-year term (April 2, 2015 to April 1, 2020). 3. Procedure 3.1 Performance Review A performance review of the information submitted by the operator in support of their renewal application was completed. This review recommended that the site be renewed based on the technical and biological assessment. This performance review is required pursuant to Subsection 72(c) of the Aquaculture Licence and Lease Regulations, and was completed on March 17, 2020. 3.2 Public Comment Period Notice of the application for the renewal of AQ#1205 for the 30-day public comment period was published on NSDFA’s website (http://novascotia.ca/fish/aquaculture/public-information/) for the period of January 9, 2020 to February 7, 2020. -

THE SAINT JOHN RIVER: a State of the Environment Report
THE SAINT JOHN RIVER: A State of the Environment Report A publication of the Canadian Rivers Institute Celebrating 10 Years of Science July 2011 The Saint John River: A State of the Environment Report ___ A publication of the Canadian Rivers Institute Celebrating 10 years of science Editors Scott D. Kidd, R. Allen Curry, Kelly R. Munkittrick The Saint John River: A State of the Environment Report 2011 Published by: Canadian River Institute University of New Brunswick Office # 202, IUC Forestry Bldg. (NF) Fredericton, New Brunswick, Canada E3B 5A3 Tel: (506) 453-4770 Fax: (506) 453-3583 E-mail: [email protected] Web: www.unb.ca/cri Reproduction of this report in part or full requires written permission from the Canadian Rivers Institute, University of New Brunswick. Library and Archives Canada Cataloguing in Publication Editors: Kidd, Scott D., 1968- Curry, R. Allen, 1960- Munkittrick, Kelly, R., 1958- The Saint John River: A State of the Environment Report ISBN 978-1-55131-158-6 1. Saint John River, New Brunswick, Canada 2. State of the environment reporting 3. Socioeconomic conditions – Saint John River Basin 4. River habitats – Saint John River 5. Water quality – Saint John River 6. Primary production – Saint John River 7. Fishes – Saint John River 8. Traditional ecological knowledge – Saint John River Basin © 2011 Canadian Rivers Institute, University of New Brunswick Acknowledgements The editors gratefully acknowledge the generous support of the New Brunswick Environmental Trust Fund in the preparation and publishing of this report. We also appreciate the time and hard work all the authors put into their chapters.