Insecticides - Development of Safer and More Effective Technologies
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pesticide Residues in Food 2007 – Report, 2007 (E)
FAO Pesticide residues PLANT PRODUCTION in food 2007 AND PROTECTION PAPER Joint FAO/WHO Meeting on Pesticide Residues 191 Report of the Joint Meeting of the FAO Panel of Experts on Pesticide Residues in Food and the Environment and the WHO Core Assessment Group on Pesticide Residues Geneva, Switzerland, 18–27 September 2007 WORLD HEALTH ORGANIZATION FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS Rome, 2007 The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations (FAO) concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned. ISBN 978-92-5-105918-0 All rights reserved. Reproduction and dissemination of material in this information product for educational or other non-commercial purposes are authorized without any prior written permission from the copyright holders provided the source is fully acknowledged. Reproduction of material in this information product for resale or other commercial purposes is prohibited without written permission of the copyright holders. Applications for such permission should be addressed to: Chief Electronic -

Historical Perspectives on Apple Production: Fruit Tree Pest Management, Regulation and New Insecticidal Chemistries
Historical Perspectives on Apple Production: Fruit Tree Pest Management, Regulation and New Insecticidal Chemistries. Peter Jentsch Extension Associate Department of Entomology Cornell University's Hudson Valley Lab 3357 Rt. 9W; PO box 727 Highland, NY 12528 email: [email protected] Phone 845-691-7151 Mobile: 845-417-7465 http://www.nysaes.cornell.edu/ent/faculty/jentsch/ 2 Historical Perspectives on Fruit Production: Fruit Tree Pest Management, Regulation and New Chemistries. by Peter Jentsch I. Historical Use of Pesticides in Apple Production Overview of Apple Production and Pest Management Prior to 1940 Synthetic Pesticide Development and Use II. Influences Changing the Pest Management Profile in Apple Production Chemical Residues in Early Insect Management Historical Chemical Regulation Recent Regulation Developments Changing Pest Management Food Quality Protection Act of 1996 The Science Behind The Methodology Pesticide Revisions – Requirements For New Registrations III. Resistance of Insect Pests to Insecticides Resistance Pest Management Strategies IV. Reduced Risk Chemistries: New Modes of Action and the Insecticide Treadmill Fermentation Microbial Products Bt’s, Abamectins, Spinosads Juvenile Hormone Analogs Formamidines, Juvenile Hormone Analogs And Mimics Insect Growth Regulators Azadirachtin, Thiadiazine Neonicotinyls Major Reduced Risk Materials: Carboxamides, Carboxylic Acid Esters, Granulosis Viruses, Diphenyloxazolines, Insecticidal Soaps, Benzoyl Urea Growth Regulators, Tetronic Acids, Oxadiazenes , Particle Films, Phenoxypyrazoles, Pyridazinones, Spinosads, Tetrazines , Organotins, Quinolines. 3 I Historical Use of Pesticides in Apple Production Overview of Apple Production and Pest Management Prior to 1940 The apple has a rather ominous origin. Its inception is framed in the biblical text regarding the genesis of mankind. The backdrop appears to be the turbulent setting of what many scholars believe to be present day Iraq. -

Mode of Action Classification
Gr oup 15: Inhibitors of chitin biosynthe si s, type 0 (Only m ajor r epr esentatives of the gr oup ar e shown) Gr oup 1: Acetylcholi ne st er a se (AChE) inhibitors (Only major representatives of the groups are shown) Mode of Action Classification Flufenoxuron No v a l u ro n Triflumuron Ac ephate Me t h a mi d o p h o s Aldic arb Ox a my l Di fl u b e n z u ron Lufenuron Teflubenzuron 15 Benz oy lur eas Me t h i o c a r b Fenitrothion Gr oup 16: Inhibitors of Gr oup 17: Moulting Gr oup 18: Ecdysone r eceptor agonists Thiodicarb Ch l o rp y ri fo s Ca rb a ry l Profenofos chitin biosynthesis, disr uptor , D ipter an type 1 Me t h o my l Ma l a t h i o n Insecticide Resistance Action Committee Pirimic arb Di meth o a te Ch ro mafe n o z i d e Ca rb o fu ra n Triazophos Me t h o x y f e n o z id e 1A Carbamates 1B Organophosphates The Key to Resistance Management Buprofez in Cy ro maz i n e 18 Diacyl- Ha l o fe n o z i d e Tebufenozide Ø Successive generations of a pest should not be treated with compounds from the same MoA Gr oup. 16 Buprofezin 17 Cyromazine hydrazines Gr oup 2: GABA-gated chlor ide channel antagonis ts Ø Not all of the cur r ent gr oupings ar e based on knowledg e of a shar ed tar get pr otein. -

Neuroactive Insecticides: Targets, Selectivity, Resistance, and Secondary Effects
EN58CH06-Casida ARI 5 December 2012 8:11 Neuroactive Insecticides: Targets, Selectivity, Resistance, and Secondary Effects John E. Casida1,∗ and Kathleen A. Durkin2 1Environmental Chemistry and Toxicology Laboratory, Department of Environmental Science, Policy, and Management, 2Molecular Graphics and Computational Facility, College of Chemistry, University of California, Berkeley, California 94720; email: [email protected], [email protected] Annu. Rev. Entomol. 2013. 58:99–117 Keywords The Annual Review of Entomology is online at acetylcholinesterase, calcium channels, GABAA receptor, nicotinic ento.annualreviews.org receptor, secondary targets, sodium channel This article’s doi: 10.1146/annurev-ento-120811-153645 Abstract Copyright c 2013 by Annual Reviews. Neuroactive insecticides are the principal means of protecting crops, people, All rights reserved livestock, and pets from pest insect attack and disease transmission. Cur- ∗ Corresponding author rently, the four major nerve targets are acetylcholinesterase for organophos- phates and methylcarbamates, the nicotinic acetylcholine receptor for neonicotinoids, the γ-aminobutyric acid receptor/chloride channel for by Public Health Information Access Project on 04/29/14. For personal use only. Annu. Rev. Entomol. 2013.58:99-117. Downloaded from www.annualreviews.org polychlorocyclohexanes and fiproles, and the voltage-gated sodium channel for pyrethroids and dichlorodiphenyltrichloroethane. Species selectivity and acquired resistance are attributable in part to structural differences in binding subsites, receptor subunit interfaces, or transmembrane regions. Additional targets are sites in the sodium channel (indoxacarb and metaflumizone), the glutamate-gated chloride channel (avermectins), the octopamine receptor (amitraz metabolite), and the calcium-activated calcium channel (diamides). Secondary toxic effects in mammals from off-target serine hydrolase inhibi- tion include organophosphate-induced delayed neuropathy and disruption of the cannabinoid system. -

EPA Method 538: Determination of Selected Organic Contaminants in Drinking Water by Aqueous Direct Injection and LC/MS/MS Summar
EPA Method 538: Determination of Selected Organic Contaminants in Drinking Water by Aqueous Direct Injection and LC/MS/MS UCT Part Numbers: SLAQ100ID21-3UM - Selectra® Aqueous C18, 100 x 2.1mm, 3µm SLAQGDC20-3UM - Selectra® Aqueous C18, Guard column, 10 x 2.0mm, 3µm SLGRDHLDR - Guard Cartridge Holder June 2015 Summary: This application outlines a direct aqueous injection-liquid chromatography/tandem mass spectrometry (DAI-LC/MS/MS) method for the determination of 11 selected organic contaminants in drinking water, including methamidophos, acephate, aldicarb sulfoxide, oxydemeton methyl, dicrotophos, aldicarb, diisopropyl methylphosphonate (DIMP), fenamiphos sulfone, fenamiphos sulfoxide, thiofanox, and quinoline [1]. Dicrotophos, oxydemeton methyl, methamidophos, and acephate are UCMR4 compounds. An Aqueous C18 HPLC column was utilized for analyte retention and separation. Calibration curves were constructed using calibration standards prepared in reagent water with preservative reagents for analyte quantitation. The responses were linear over the entire analytical ranges (R2 ≥ 0.9970). Excellent accuracy (90 - 111%) and precision (RSD% < 20%, n=7) were achieved for fortified reagent water and tap water samples. Procedure: 1. Preserve drinking water sample with 64 mg/L of sodium omadine (antimicrobial) and 1.5 g/L of ammonium acetate (binding free chlorine). 2. Mix 0.99 mL of the preserved water sample with 10 μL of 0.4-12.5 ng/μL internal standard mixture, and vortex for 30 sec. 3. Inject 50 μL onto LC/MS/MS equipped with an aqueous -

Order DIPTERA. Family HIPPOBOSCIDAE
781 Genus Rhinolophopsylla. ! • ' . ~ • Rhin~lophopsylla. l . RhinolOphopsylla capensis ·Jordan and Rothschild. Rhinolophopsylla capensis Jord. and Rothsch, Ectoparasites, I, Pt. 3, .p. 148, f. 126-128 (1921). Described from a small series of oo and ~~ taken off Nycteris cape'f'Sis , (pape, long-eared ba,t) at . Mfongosi, Zululand. Order DIPTERA. Family HIPPOBOSCIDAE. , Th~ .flies 'incluq.ed in this family. are pa,rasitic on mammals and birds. TABLE OF SouTH Aii~ICAN GENERA (after Speiser). 1. Wings well developed and function:;tl. 2 Wings rudimentary or wanting ..... , .. :.. 8 2. Claws with the usual two points (heel and tip), ·parasitic on mammals.. 3 Claws with three teeth ; parasitic on birds,........ , : . .. 5 3. Head of normal form, not broadly impinging on the thorax, freely movable ; ocelli absent; wings always present ...... Hippobosca. Head :flat, broadly impinging on the thorax ; wings usually becoming · detached, especially in, the females, leaving only a shred. 4 ·1·. Ocelli absent . ........ ......... ................. ... Echestypus. Ocelli' presen£ .. : .........· ... ·........................... Lipoptena. 5. Ocelli present ; anal cell present ............ <· ........ Ornithomyia. Ocelli absent; anal cell absent........ 6 6. Wings of usual shape ; scutellum not truncate. 7 Wings lanceolate, rounded at tip; scutellum truncate ... ... Lynchia. 7. Distance of oral borders from frontal suture distinctly less than from the suture to the vertex ....... ........ .... .. ........ Olfersia. Distance from oral borders to suture -

Chemical Name Federal P Code CAS Registry Number Acutely
Acutely / Extremely Hazardous Waste List Federal P CAS Registry Acutely / Extremely Chemical Name Code Number Hazardous 4,7-Methano-1H-indene, 1,4,5,6,7,8,8-heptachloro-3a,4,7,7a-tetrahydro- P059 76-44-8 Acutely Hazardous 6,9-Methano-2,4,3-benzodioxathiepin, 6,7,8,9,10,10- hexachloro-1,5,5a,6,9,9a-hexahydro-, 3-oxide P050 115-29-7 Acutely Hazardous Methanimidamide, N,N-dimethyl-N'-[2-methyl-4-[[(methylamino)carbonyl]oxy]phenyl]- P197 17702-57-7 Acutely Hazardous 1-(o-Chlorophenyl)thiourea P026 5344-82-1 Acutely Hazardous 1-(o-Chlorophenyl)thiourea 5344-82-1 Extremely Hazardous 1,1,1-Trichloro-2, -bis(p-methoxyphenyl)ethane Extremely Hazardous 1,1a,2,2,3,3a,4,5,5,5a,5b,6-Dodecachlorooctahydro-1,3,4-metheno-1H-cyclobuta (cd) pentalene, Dechlorane Extremely Hazardous 1,1a,3,3a,4,5,5,5a,5b,6-Decachloro--octahydro-1,2,4-metheno-2H-cyclobuta (cd) pentalen-2- one, chlorecone Extremely Hazardous 1,1-Dimethylhydrazine 57-14-7 Extremely Hazardous 1,2,3,4,10,10-Hexachloro-6,7-epoxy-1,4,4,4a,5,6,7,8,8a-octahydro-1,4-endo-endo-5,8- dimethanonaph-thalene Extremely Hazardous 1,2,3-Propanetriol, trinitrate P081 55-63-0 Acutely Hazardous 1,2,3-Propanetriol, trinitrate 55-63-0 Extremely Hazardous 1,2,4,5,6,7,8,8-Octachloro-4,7-methano-3a,4,7,7a-tetra- hydro- indane Extremely Hazardous 1,2-Benzenediol, 4-[1-hydroxy-2-(methylamino)ethyl]- 51-43-4 Extremely Hazardous 1,2-Benzenediol, 4-[1-hydroxy-2-(methylamino)ethyl]-, P042 51-43-4 Acutely Hazardous 1,2-Dibromo-3-chloropropane 96-12-8 Extremely Hazardous 1,2-Propylenimine P067 75-55-8 Acutely Hazardous 1,2-Propylenimine 75-55-8 Extremely Hazardous 1,3,4,5,6,7,8,8-Octachloro-1,3,3a,4,7,7a-hexahydro-4,7-methanoisobenzofuran Extremely Hazardous 1,3-Dithiolane-2-carboxaldehyde, 2,4-dimethyl-, O- [(methylamino)-carbonyl]oxime 26419-73-8 Extremely Hazardous 1,3-Dithiolane-2-carboxaldehyde, 2,4-dimethyl-, O- [(methylamino)-carbonyl]oxime. -
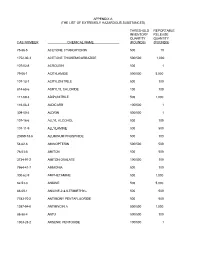
The List of Extremely Hazardous Substances)
APPENDIX A (THE LIST OF EXTREMELY HAZARDOUS SUBSTANCES) THRESHOLD REPORTABLE INVENTORY RELEASE QUANTITY QUANTITY CAS NUMBER CHEMICAL NAME (POUNDS) (POUNDS) 75-86-5 ACETONE CYANOHYDRIN 500 10 1752-30-3 ACETONE THIOSEMICARBAZIDE 500/500 1,000 107-02-8 ACROLEIN 500 1 79-06-1 ACRYLAMIDE 500/500 5,000 107-13-1 ACRYLONITRILE 500 100 814-68-6 ACRYLYL CHLORIDE 100 100 111-69-3 ADIPONITRILE 500 1,000 116-06-3 ALDICARB 100/500 1 309-00-2 ALDRIN 500/500 1 107-18-6 ALLYL ALCOHOL 500 100 107-11-9 ALLYLAMINE 500 500 20859-73-8 ALUMINUM PHOSPHIDE 500 100 54-62-6 AMINOPTERIN 500/500 500 78-53-5 AMITON 500 500 3734-97-2 AMITON OXALATE 100/500 100 7664-41-7 AMMONIA 500 100 300-62-9 AMPHETAMINE 500 1,000 62-53-3 ANILINE 500 5,000 88-05-1 ANILINE,2,4,6-TRIMETHYL- 500 500 7783-70-2 ANTIMONY PENTAFLUORIDE 500 500 1397-94-0 ANTIMYCIN A 500/500 1,000 86-88-4 ANTU 500/500 100 1303-28-2 ARSENIC PENTOXIDE 100/500 1 THRESHOLD REPORTABLE INVENTORY RELEASE QUANTITY QUANTITY CAS NUMBER CHEMICAL NAME (POUNDS) (POUNDS) 1327-53-3 ARSENOUS OXIDE 100/500 1 7784-34-1 ARSENOUS TRICHLORIDE 500 1 7784-42-1 ARSINE 100 100 2642-71-9 AZINPHOS-ETHYL 100/500 100 86-50-0 AZINPHOS-METHYL 10/500 1 98-87-3 BENZAL CHLORIDE 500 5,000 98-16-8 BENZENAMINE, 3-(TRIFLUOROMETHYL)- 500 500 100-14-1 BENZENE, 1-(CHLOROMETHYL)-4-NITRO- 500/500 500 98-05-5 BENZENEARSONIC ACID 10/500 10 3615-21-2 BENZIMIDAZOLE, 4,5-DICHLORO-2-(TRI- 500/500 500 FLUOROMETHYL)- 98-07-7 BENZOTRICHLORIDE 100 10 100-44-7 BENZYL CHLORIDE 500 100 140-29-4 BENZYL CYANIDE 500 500 15271-41-7 BICYCLO[2.2.1]HEPTANE-2-CARBONITRILE,5- -

The Pesticides Are Generally Classified Into Various Groups Based
Lecture 19 PESTICIDES GROUPS Groups of pesticides : The pesticides are generally classified into various groups based on pest organism against which the compounds are used, their chemical nature, mode of entry and mode of action. 1. Based on organisms a) Insecticides : Chemicals used to kill or control insects (eg.) endosulfan, malathion b) Rodenticides : Chemicals exclusively used to control rats (eg.) Zinc phosphide c) Acaricides : Chemicals used to control mites on crops / animals (eg.) Dicofol d) Avicides : Chemicals used to repel the birds (eg.) Anthraquionone e) Molluscicides : Chemicals used to kill the snails and slugs (eg.) Metaldehyde f) Nematicides : Chemicals used to control nematodes (eg.) Ethylene dibromide g) Fungicides : Chemicals used to control plant diseases caused by fungi (eg.) Copper oxy cholirde h) Bactericide : Chemicals used to control the plant diseases caused by bacteria (eg.) Streptomycin sulphate i) Herbicide : Chemicals used to control weeds (eg.) 2,4, - D 2. Based on mode of entry a) Stomach poison : The insecticide applied in the leaves and other parts of the plant when ingested, act in the digestive system of the insect and bring about kill (eg.) Malathion. b) Contact Poison : The toxicant which brings about death of the pest species by means of contact (eg.) Fenvalerate. c) Fumigant : Toxicant enter in vapour form into the tracheal system (respiratory poison) through spiracles (eg.) Aluminium phosphide d) Systemic poison : Chemicals when applied to plant or soil are absorbed by foliage (or) roots and translocated through vascular system and cause death of insect feeding on plant. (eg.) Dimethoate. 3. Based on mode of action a) Physical poison : Toxicant which brings about kill of one insect by exerting a physical effect (eg.) Activated clay. -

Code Chemical P026 1-(O-Chlorophenyl)Thiourea P081 1
Code Chemical P026 1-(o-Chlorophenyl)thiourea P081 1,2,3-Propanetriol, trinitrate (R) P042 1,2-Benzenediol, 4-[1-hydroxy-2-(methylamino)ethyl]-, (R)- P067 1,2-Propylenimine P185 1,3-Dithiolane-2-carboxaldehyde, 2,4-dimethyl-, O- [(methylamino)- carbonyl]oxime 1,4,5,8-Dimethanonaphthalene, 1,2,3,4,10,10-hexa- chloro-1,4,4a,5,8,8a,-hexahydro-, P004 (1alpha,4alpha, 4abeta,5alpha,8alpha,8abeta)- 1,4,5,8-Dimethanonaphthalene, 1,2,3,4,10,10-hexa- chloro-1,4,4a,5,8,8a-hexahydro-, P060 (1alpha,4alpha, 4abeta,5beta,8beta,8abeta)- P002 1-Acetyl-2-thiourea P048 2,4-Dinitrophenol P051 2,7:3,6-Dimethanonaphth [2,3-b]oxirene, 3,4,5,6,9,9 -hexachloro-1a,2,2a,3,6,6a,7,7a- octahydro-, (1aalpha,2beta,2abeta,3alpha,6alpha,6abeta,7 beta, 7aalpha)-, & metabolites 2,7:3,6-Dimethanonaphth[2,3-b]oxirene, 3,4,5,6,9,9- hexachloro-1a,2,2a,3,6,6a,7,7a- P037 octahydro-, (1aalpha,2beta,2aalpha,3beta,6beta,6aalpha,7 beta, 7aalpha)- P045 2-Butanone, 3,3-dimethyl-1-(methylthio)-, O-[methylamino)carbonyl] oxime P034 2-Cyclohexyl-4,6-dinitrophenol 2H-1-Benzopyran-2-one, 4-hydroxy-3-(3-oxo-1- phenylbutyl)-, & salts, when present at P001 concentrations greater than 0.3% P069 2-Methyllactonitrile P017 2-Propanone, 1-bromo- P005 2-Propen-1-ol P003 2-Propenal P102 2-Propyn-1-ol P007 3(2H)-Isoxazolone, 5-(aminomethyl)- P027 3-Chloropropionitrile P047 4,6-Dinitro-o-cresol, & salts P059 4,7-Methano-1H-indene, 1,4,5,6,7,8,8-heptachloro- 3a,4,7,7a-tetrahydro- P008 4-Aminopyridine P008 4-Pyridinamine P007 5-(Aminomethyl)-3-isoxazolol 6,9-Methano-2,4,3-benzodioxathiepin, 6,7,8,9,10,10- -
![Ectoparasites of Feral Horses [Equus Ferus Caballus (Linnaeus., 1758)] on Karadag˘ Mountain, Karaman, Turkey](https://docslib.b-cdn.net/cover/2536/ectoparasites-of-feral-horses-equus-ferus-caballus-linnaeus-1758-on-karadag-mountain-karaman-turkey-1062536.webp)
Ectoparasites of Feral Horses [Equus Ferus Caballus (Linnaeus., 1758)] on Karadag˘ Mountain, Karaman, Turkey
J Parasit Dis https://doi.org/10.1007/s12639-020-01234-4 ORIGINAL ARTICLE Ectoparasites of feral horses [Equus ferus caballus (Linnaeus., 1758)] on Karadag˘ Mountain, Karaman, Turkey 1 1 1 2 Bilal Dik • Onur Ceylan • Ceylan Ceylan • Mustafa Agah Tekindal • 3 2 1 Asma Semassel • Gonca So¨nmez • O¨ zlem Derinbay Ekici Received: 20 February 2020 / Accepted: 3 June 2020 Ó Indian Society for Parasitology 2020 Abstract Approximately 250 feral horses [Equus ferus Keywords Tick Á Louse Á Bovicola equi Á caballus (Linnaeus, 1758)] living on Karadag˘ Mountain Hippobosca equina Á Haemaphysalis parva near Karaman City were caught by Kazakh horse herdsmen with permission of the Turkish Ministry of Agriculture and Forestry and brought to a farm in Karkın village in Konya Introduction Province, 70 km from Karadag˘, in November, 2017. This study was carried out to determine the presence of Linnaeus described domestic horse as Equus caballus in ectoparasites infesting a subsample of 36 feral horses. The 1758. Equus caballus, which is known as Equus ferus, horses were visually inspected, and then their bodies were contains seven subspecies some of which are extinct checked by hand for ectoparasites. Thirty-five (97.2%) (Bennett and Hoffmann 1999). The origin of domestic were infested with at least one of five species of ectopar- horses (Equus caballus caballus and Equus ferus caballus) asites: Bovicola equi (Linnaeus, 1758), Hippobosca equina stems from potential wild ancestors known as the Prze- (Linnaeus, 1758), Haemaphysalis parva (Neuman, 1897), walski horse and the Tarpan horse. Even today, the Prze- Hyalomma excavatum (Koch, 18449), Dermacentor walski horse (Equus ferus przewalskii Poliakov, 1881) lives marginatus (Sulzer, 1776). -

Bark Beetles Integrated Pest Management for Home Gardeners and Landscape Professionals
BARK BEETLES Integrated Pest Management for Home Gardeners and Landscape Professionals Bark beetles, family Scolytidae, are California now has 20 invasive spe- common pests of conifers (such as cies of bark beetles, of which 10 spe- pines) and some attack broadleaf trees. cies have been discovered since 2002. Over 600 species occur in the United The biology of these new invaders is States and Canada with approximately poorly understood. For more informa- 200 in California alone. The most com- tion on these new species, including mon species infesting pines in urban illustrations to help you identify them, (actual size) landscapes and at the wildland-urban see the USDA Forest Service pamphlet, interface in California are the engraver Invasive Bark Beetles, in References. beetles, the red turpentine beetle, and the western pine beetle (See Table 1 Other common wood-boring pests in Figure 1. Adult western pine beetle. for scientific names). In high elevation landscape trees and shrubs include landscapes, such as the Tahoe Basin clearwing moths, roundheaded area or the San Bernardino Mountains, borers, and flatheaded borers. Cer- the Jeffrey pine beetle and mountain tain wood borers survive the milling Identifying Bark Beetles by their Damage pine beetle are also frequent pests process and may emerge from wood and Signs. The species of tree attacked of pines. Two recently invasive spe- in structures or furniture including and the location of damage on the tree cies, the Mediterranean pine engraver some roundheaded and flatheaded help in identifying the bark beetle spe- and the redhaired pine bark beetle, borers and woodwasps. Others colo- cies present (Table 1).