Redalyc.Palaeoenvironmental, Palaeoecological And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Palaeoenvironmental, Palaeoecological and Palaeobiogeographical Implications of Mixed Fresh Water and Brackish Marine Assemblage
REVISTAKhosla MEXICANA DE CIENCIAS GEOLÓGICAS v. 32, núm. 2, 2015, p. 344-357 Palaeoenvironmental, palaeoecological and palaeobiogeographical implications of mixed fresh water and brackish marine assemblages from the Cretaceous-Palaeogene Deccan intertrappean beds at Jhilmili, Chhindwara District, central India Ashu Khosla Centre of Advanced Study in Geology, Panjab University, Sector-14, Chandigarh-160014, India. [email protected] ABSTRACT depósitos terrestres y fluviales-lacustres. La presencia de foraminíferos planctónicos y ostrácodos de agua salobre en las capas Jhilmili del The Deccan infra- and intertrappeans volcano-sedimentary Daniano inferior intercaladas en los “traps”, descritas con anterioridad, sequences have been considered as terrestrial to fluvio-lacustrine indica paleoambiente, paleoecología y paleobiogeografía complejos. deposits. Presence of planktonic foraminifera and brackish water Los sedimentos intercalados en los “traps” tienen 14 m de espesor y ostracods in the Jhilmili basal Danian intertrappean beds, recorded se encuentran entre dos flujos basálticos del Deccan; los sedimentos earlier, indicates complex palaeoenvironment, palaeoecology and se depositaron en condiciones terrestres, palustres y de planicies de palaeobiogeography. The intertrappean sediments span 14 m between inundación. En medio de esta sección se encuentra una capa con espesor two Deccan basaltic flows deposited under terrestrial, palustrine and de 60 cm (unidad 3) que contiene ostrácodos de agua dulce, carofitas y flood plain environments. In the middle of this section is a 60 cm- foraminíferos planctónicos del Daniano temprano. Paleoecológicamente, thick layer (unit 3) that contains fresh water ostracods, charophytes el conjunto de ostrácodos incluye nadadores activos (Cypridopsis, and Early Danian planktonic foraminifera. Palaeoecologically, the Cypria, Mongolianella, Paracypretta y Zonocypris) y nadadores pobres ostracod assemblage includes active swimmers (Cypridopsis, Cypria, (Limnocythere, Gomphocythere, Frambocythere y Darwinula). -

Madhya Pradesh)
CENSUS OF INDIA, 1951 (Madhya Pradesh) JABALPUR District Census Handbook By J. D. KERAWALLA, M.Sc. 01 the Indian Administrative Service .'>''Jpzrintendent of Census Operations, Madhya Pradesh NAGPUR GoVERNMENT PRINTING, MADHYA PRADESH CONTENTS PAGE htroduction i-iii A.-General Population Tables- I. Area, Houses and Population I II. Variation in Population during fifty years 1 III. Towns and Villages classified by Population 2 IV. Towns classified by Population with Variation since 1901 3 V. Towns arranged territorially with Population by Livelihood Classes - 4 B.-Economic Tables- I. Livelihood Classes and Sub-Classes - 5-10 .. {For Total Population 11-28 II. Seco~dary Means of Llvehhood For Displaced Persons 29-46 III. Employers, Employees and Independent Workers by Industries and Services, 47-93 Divisions and Sub-Divisions. The District Index of Non-Agricultural Occupations - 94-91' C.-Household and Age (Sample) Tables- 1. Household (Size and Composition) - 98 . {For Sample Population- 99-103 II. Llvehhood Classes by Age Groups For Displaced Persons _ 104-109 III. Age and Civil Condition 110-116 IV. Age and Literacy 117-122 . {For Sample Population - 123-125 V. SIngle Year Age Returns F D' I d P or ISP ace ersons 126-128 D.-Social and Cultural Tables I. (i) Mother tongue 129--131 I. (ii) Bilingualism 132~133 II. Religion 134 III. Scheduled Castes, Scheduled Tribes and Anglo-Indians 135 IV. Migrants 135-141 V. Displaced Persons by District of Origin and Year of Arrival in India 142-143 VI. Non-Indian Nationals 144 VII. Livelihood Classe(by Educational-Standards 145-153 E.-Summary Figures for the District and Tahsils 154-155 The Primary Census Abstract (showing Distribution of the General Population by Livelihood 156-215 Classes and Literacy, and Number of occupied houses in Villages and Wards). -

Statistical Report General Election, 1998 The
STATISTICAL REPORT ON GENERAL ELECTION, 1998 TO THE LEGISLATIVE ASSEMBLY OF MADHYA PRADESH ELECTION COMMISSION OF INDIA NEW DELHI Election Commission of India – State Elections, 1998 Legislative Assembly of Madhya Pradesh STATISCAL REPORT ( National and State Abstracts & Detailed Results) CONTENTS SUBJECT Page No. Part – I 1. List of Participating Political Parties 1 - 2 2. Other Abbreviations And Description 3 3. Highlights 4 4. List of Successful Candidates 5 - 12 5. Performance of Political Parties 13 - 14 6. Candidate Data Summary 15 7. Electors Data Summary 16 8. Women Candidates 17 - 25 9. Constituency Data Summary 26 - 345 10. Detailed Results 346 - 413 Election Commission of India-State Elections, 1998 to the Legislative Assembly of MADHYA PRADESH LIST OF PARTICIPATING POLITICAL PARTIES PARTYTYPE ABBREVIATION PARTY NATIONAL PARTIES 1 . BJP Bharatiya Janata Party 2 . BSP Bahujan Samaj Party 3 . CPI Communist Party of India 4 . CPM Communist Party of India (Marxist) 5 . INC Indian National Congress 6 . JD Janata Dal (Not to be used in General Elections, 1999) 7 . SAP Samata Party STATE PARTIES 8 . ICS Indian Congress (Socialist) 9 . INLD Indian National Lok Dal 10 . JP Janata Party 11 . LS Lok Shakti 12 . RJD Rashtriya Janata Dal 13 . RPI Republican Party of India 14 . SHS Shivsena 15 . SJP(R) Samajwadi Janata Party (Rashtriya) 16 . SP Samajwadi Party REGISTERED(Unrecognised ) PARTIES 17 . ABHM Akhil Bharat Hindu Mahasabha 18 . ABJS Akhil Bharatiya Jan Sangh 19 . ABLTC Akhil Bhartiya Lok Tantrik Congress 20 . ABMSD Akhil Bartiya Manav Seva Dal 21 . AD Apna Dal 22 . AJBP Ajeya Bharat Party 23 . BKD(J) Bahujan Kranti Dal (Jai) 24 . -

THE URBAN LAND (CEILING and REGULATION) ACT, 1976 No
THE URBAN LAND (CEILING AND REGULATION) ACT, 1976 No. 33 OF 1976 [17th February, 1976] An Act to provide for the imposition of a ceiling on vacant land in urban agglomerations, for the acquisition of such land in excess of the ceiling limit, to regulate the construction of buildings on such land and for matters connected therewith, with a view to preventing the concentration of urban land in the hands of a few persons and speculation and profiteering therein and with a view to bringing about an equitable distri- bution of land in urban agglomerations to subserve the common good. WHEREAS it is expedient to provide for the imposition of a ceiling on vacant land in urban agglomerations , for the acquisition of such land in excess of the ceiling limit, to regulate the construction of buildings on such land and for matters connected therewith , with a view to prevent- ing the concentration of urban land in the hands of a few persons and speculation and profiteering therein and with a view to bringing about an equitable distribution of land in urban agglomerations to subserve the common good; AND WHEREAS Parliament has, no power to lna$e laws for the States with respect to the matters aforesaid except as provided in articles 249 and 250 of the Constitution; 2 Urban Land (Ceiling and Regulation) LACT 33 AND WHEREAS in pursuance of clause (1) of article 252 of the Consti- tution resolutions have been passed by all the Houses of the Legislatures of the States of Andhra Pradesh, Gujarat, Haryana, Himachal Pradesh, Karnataka, Maharashtra, Orissa, Punjab, Tripura, Uttar Pradesh and West Bengal that the matters aforesaid should be regulated in those States by Parliament by law; BE it enacted by Parliament in the Twenty-seventh Year of the Republic of India as follows:- CHAPTER I PRELIMINARY Short 1. -

Madhya Pradesh
CENSUS OF INDIA 1971 SERIES 10 MADHYA PRADESH PART VI-A A. K. PANDYA Of the Indian Administrative Service DIRECTOR OF CENSUS OPERATIONS MADHYA PRADESH 0-e.. J:~ (J)wl[) «..Jo~ 0:(.) LLJ Cl.N ~~J:CO (j) Q z « 3 1: o.... o <C a: V) w t1> o Z ~ o 3: en G\'" o ~ 0() I-O .. '"I o o g. 0 u. 0 o o ): o o 0 p o o 6 o 0 oJ o ill z ~ N 0 '" o PRG. 5.71 (MP) (N) 500 PRINTED BY THE MANAGER, GOVERNMENT OF INDIA PRESS, NASIK-422006 AND PUBLlSHED BY THE CONTROLLER OF PUBLICATIONS DELHI-llOOO6 1975 1971 CENSUS PUBLICATIONS, MADHYA PRADESH (All the Census PubLicatwns of this State will bear series No. 10) PART I Census General Report including Subsidiary (in Suf>.Parts) Tables. Census Tables on population. PART II-A PART U-B Eco~omic Tables. (in Suf>.Parts) PART H-C Social and Cultural Tables. (in Sub-Parts) PART III-A, Eitablishment Report and Subsidiary Tables. PART III-B Establishment Tables. PART IV Housing Report and Tables. PART V Special Tables and Ethnographic Notes on (in Suf>.Parts) Scheduled Castes and Scheduled Tribes. PART VI-A Town Directory. PART VI-B Special Survey Reports On selected towns. PART VI-C _.. ' Survey Reports on selected villages. PART VII Special Report on Graduates and Technical Personnel. PART VHJ-A Administration RepoFt-Enumeration. PART VHJ-B -\dministration Report-Tabulation. PART IX Census Atlas. FART lX-A A.dministrative Atlas. STATE GOVERNMENT PUBLICATIONS PART X-A Village and Town Directory_ PART X-B Village and TOWn Primary Census Abstrads. -

District Census Handbook, Jabalpur, Part XIII-A, Series-11
~. ~T ~, "'~)lI' ~;rcr; ~, f; , ll~, ~11r.n, ~~w. I 1981 CENSUS-PUBLICATION p~ t IfB I Census Pujli~alions. Series 1J in All India Series will be published in the folJowing par"! GOVERNMENT OF INDIA PUBLICATIONS Part 1- Administration Report-Enumeration Part I-B Administration Report-Tabulation Part II-A General Population Tables Part II-B Primary Census Abstract Part III Geaeral Economic Tables Part IV Social and Cultural Tables Part V Migration Tables :Part VI Fertility Tables Part VII Tables on Houses and Disabled 'Population ' Part VIII Household Tables Part IX Special Tables on Scheduled Castes and Scheduled Tribes Part X-A Town Directory Part X-,B Survey Reports on selected Towns , . Part X-C Survey Reports ~!l seJecte~ ViHages Part XI Ethnographic Notes and special studies 011 Sche<ftultit Castes and Scheduled Tribes Part XU Census Atlas Paper 1 of 1982 Primary Census Abstract for Scheduled Castes and Scheduled Tribe. Paper 1 of 1984 • Household Population by Religion of Head of Household STATE GOVERNMENT PURLTCAT ONS Part XIII-A and B District Census Handbook for each of the 4S distrcts in the State (Village and Town Directory and Primary Census Abstract) ,~~~) '" CONTENTS, ~'S 'ages 1 '%t'f~~ Porewotd I-IV 2 sr~"T'!'~r Preface V-VI 3 r~~t;r~r DJstrict Map 4 ~~T'i artCfi~ Jmportant Statistics VIf 5 f~~lSfo(f~$ f~) Anal)tical Note o~TilTT(1f'" gcq1Jf\' ; at~~f'i{ij' ;jfTf(f 11'1:: 81'iW~ Note & Explanatwn'\;. List of Scheduled Castes and Scheduled Tn"bec; Order \iA'mm ot\" ~) ( ftlu"') , Mlttlli 1976; (Amendment) Act, 1976, History and f\iA'T 'lfiflTlJfifT ,!f~ 'liT ~~~ atR el"l' I Scope of District Census Handbook, ~lJfl~~'1l fic'tOft Analytical Note XXI-XXXVIII 6 (\'~~iT~l $ ;I:f am Tabfil Mal'9 7 Ws $-VTlf f;Rm~' Section I-VILLAGE DIRECTORY 1-413 (1). -

NPS Authorised Branches
Union Bank of India Authorized Branches for National Pension System (NPS) S.No. REGION OFFICE Sol ID Branch Branch Code 1 Ahmedabad 50350 AHMEDABAD - IFB 550353 2 Ahmedabad 50360 AHMEDABAD - SSI - FINANCE BRANCH 550361 3 Ahmedabad 40660 ARNEJ 540668 4 Ahmedabad 35350 ASARWA - AHMEDABAD 535354 5 Ahmedabad 37200 ASHRAM ROAD - AHMEDABAD 537209 6 Ahmedabad 44360 BAGODARA 544361 7 Ahmedabad 45480 BAPUNAGAR - AHMEDABAD 545481 8 Ahmedabad 31360 BAREJA 531367 9 Ahmedabad 39180 BHAIRAVNATH ROAD - AHMEDABAD 539180 10 Ahmedabad 53810 BODAKDEV - AHMEDABAD 553816 11 Ahmedabad 74610 BOPAL 574619 12 Ahmedabad 43550 C.G. ROAD - AHMEDABAD 543551 13 Ahmedabad 58180 CHANDKHEDA - AHMEDABAD 558184 14 Ahmedabad 71500 CHANGODAR 571504 15 Ahmedabad 58480 CIVIL HOSP. COMPOUND 558486 16 Ahmedabad 31260 DHANLAXMI MARKET - AHMEDABAD 531260 17 Ahmedabad 59560 DHOLKA 559563 18 Ahmedabad 49140 DR. JIVRAJ MEHTA ROAD - AHMEDABAD 549142 19 Ahmedabad 44910 DR. S. R. MARG - AHMEDABAD 544914 20 Ahmedabad 55790 DRIVE IN ROAD - AHMEDABAD 555797 21 Ahmedabad 31270 DUDHESHWAR ROAD - AHMEDABAD 531278 22 Ahmedabad 77300 DURGI 577308 23 Ahmedabad 31280 ELLIS BRIDGE - AHMEDABAD 531286 24 Ahmedabad 31290 GANDHI ROAD - AHMEDABAD 531294 25 Ahmedabad 74620 GIDC VATVA - AHMEDABAD 574627 26 Ahmedabad 31300 GOMTIPUR - AHMEDABAD-SSI 531308 27 Ahmedabad 57540 ISANPUR 557544 28 Ahmedabad 36190 JAMALPUR - AHMEDABAD 536199 29 Ahmedabad 42230 JODHPUR TEKRA - AHMEDABAD 542237 30 Ahmedabad 40340 KATHWADA 540340 31 Ahmedabad 36150 KHANPUR - AHMEDABAD 536156 32 Ahmedabad 64740 KUJAD 564745 33 Ahmedabad -
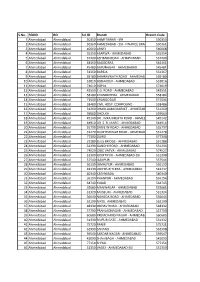
S.No. FGMO RO Sol ID Branch Branch Code 1 Ahmedabad Ahmedabad
S.No. FGMO RO Sol ID Branch Branch Code 1 Ahmedabad Ahmedabad 50350 AHMEDABAD - IFB 550353 2 Ahmedabad Ahmedabad 50360 AHMEDABAD - SSI - FINANCE BRANCH 550361 3 Ahmedabad Ahmedabad 40660 ARNEJ 540668 4 Ahmedabad Ahmedabad 35350 ASARWA - AHMEDABAD 535354 5 Ahmedabad Ahmedabad 37200 ASHRAM ROAD - AHMEDABAD 537209 6 Ahmedabad Ahmedabad 44360 BAGODARA 544361 7 Ahmedabad Ahmedabad 45480 BAPUNAGAR - AHMEDABAD 545481 8 Ahmedabad Ahmedabad 31360 BAREJA 531367 9 Ahmedabad Ahmedabad 39180 BHAIRAVNATH ROAD - AHMEDABAD 539180 10 Ahmedabad Ahmedabad 53810 BODAKDEV - AHMEDABAD 553816 11 Ahmedabad Ahmedabad 74610 BOPAL 574619 12 Ahmedabad Ahmedabad 43550 C.G. ROAD - AHMEDABAD 543551 13 Ahmedabad Ahmedabad 58180 CHANDKHEDA - AHMEDABAD 558184 14 Ahmedabad Ahmedabad 71500 CHANGODAR 571504 15 Ahmedabad Ahmedabad 58480 CIVIL HOSP. COMPOUND 558486 16 Ahmedabad Ahmedabad 31260 DHANLAXMI MARKET - AHMEDABAD 531260 17 Ahmedabad Ahmedabad 59560 DHOLKA 559563 18 Ahmedabad Ahmedabad 49140 DR. JIVRAJ MEHTA ROAD - AHMEDABAD 549142 19 Ahmedabad Ahmedabad 44910 DR. S. R. MARG - AHMEDABAD 544914 20 Ahmedabad Ahmedabad 55790 DRIVE IN ROAD - AHMEDABAD 555797 21 Ahmedabad Ahmedabad 31270 DUDHESHWAR ROAD - AHMEDABAD 531278 22 Ahmedabad Ahmedabad 77300 DURGI 577308 23 Ahmedabad Ahmedabad 31280 ELLIS BRIDGE - AHMEDABAD 531286 24 Ahmedabad Ahmedabad 31290 GANDHI ROAD - AHMEDABAD 531294 25 Ahmedabad Ahmedabad 74620 GIDC VATVA - AHMEDABAD 574627 26 Ahmedabad Ahmedabad 31300 GOMTIPUR - AHMEDABAD-SSI 531308 27 Ahmedabad Ahmedabad 57540 ISANPUR 557544 28 Ahmedabad Ahmedabad 36190 -

The Urban Land (Ceiling and Regulation) Act, 1976
CHAPTER I Preliminary 1. Short title, application and commencement. CHAPTER II Definitions 2. Definitions. CHAPTER III Ceiling on vacant land 3. Persons not entitled to hold vacant land in excess of the ceiling limit. 4. Ceiling limit. 5. Transfer of vacant land. 6. Persons holding vacant land in excess of ceiling limit to file statement. 7. Filing of statement in cases where vacant land held by a person is situated within the jurisdiction of two or more competent authorities. 8. Preparation of draft statement as regards vacant land held in excess of ceiling limit. 9. Final Statement. 10. Acquisition of vacant land in excess of ceiling limit. 11. Payment of amount for vacant land acquired. 12. Constitution of Urban Land Tribunal and appeal to Urban Land Tribunal. 13. Second appeal to High Court. 14. Mode of payment of amount. 15. Ceiling limit on future acquisition by inheritance, bequest or by sale in execution of decrees, etc. 16. Certain persons to file statements when the Act is adopted subsequently by any State. 17. Power to enter upon any vacant land. 18. Penalty for concealment, etc., of particulars of vacant land. 19. Chapter not to apply to certain vacant lands. 20. Power to exempt. 21. Excess vacant land not to be treated as excess in certain cases. 22. Retention of vacant land under certain circumstances. 23. Disposal of vacant land acquired under the Act. 24. Special provisions regarding disposal of vacant land in favour of certain persons. CHAPTER IV Regulation of transfer and use of urban property 25. Definition. 26. Notice to be given before transfer of vacant land. -

General Population Tables and Summary Figures for Districts, Part
Census of India, 1951 VOLUME VII MADHYA PRADESH PART II-A General Population Tables and Summary Figures for Districts By J. D. KERA WALLA, . M. Se. Of the Indian Administrati'l.. e Service SUperintendent of Census Operations, Madhya Pradesh NAGPUR GoVERNMENT PRINTINC, MADHYA PRADESH 1952 NOTE 1. Census of India, 1951, Volume VII, for lVladhya Pradesh, IS divided into the following parts :- PART I Report. PART II Tables in four volumes as follows:- Part II-A General Population Tables of the "A" Series and Summary Figures for Districts constituting the "E" .series. Part II-B The Economic Tables of the " B" Series. Part Il-C Household and Age (Sample) Tables of the « C" Series and Social and Cultural Tables of the " D" Series. Part II-D The Fertility Tables of the " F" Series_. 2. The Administration Report is in two parts- PART I Enumeration (The Enumeration Procedure-Training of Staff-The Census Questionnaire and details of putting it across to the People-The National Register of Citizens-Verification of the 1951 Census Count and the Sample Census Enquiry, etc.). PART II Tabulation. CONTENTS Table Pages A.-General Population Tables A-I Area, Houses and Population to 3 A-II Variation in Population during Fifty Years 5 to 9 A-III Towns and Villages classified by Population 11 -to 15 A-IV Towns classified by Population with Variation since 1901 17 to 36 A-V Towns arranged territorially with Population by Livelihood Classes 37 to 47 E.-Summary Figures for Districts E Summary Figures by Districts and Tahsils 49 to 88 TABLE A-I Area, houses and ·population This table shows for each district in the State the area, number of towns and villages, number of occupied houses and total urban and rural population, male and female. -

Administration Report A-Enumeration, Part VIII, Vol-VIII
CENSUS OF INDIA, 1961 VOLUME VIII M.ADHYA PRADESH PART VIII ADMINIST.RATION REPORT A-ENUMERATION G. JAG~THPATHI Superintendent if Census Operations, Madhya Pradesh BHOPAL GoVERNMENT CENTRAL PREss 1963 MADHYA PRADESH 1961, Census Publications The Madhya Pradesh State 1961 Census Publications, which will form Volume VIn in All-India Census Series, called "Census of India, 1961", will be in the following parts :-:-- PART lA-General Report. PART IE-Report on Vital Statistics. PART IC-Subsidiary Tables including reprints, if any, from previous Census Reports. PART II A-General Population Tables (A Series) for the State and Primary Census Abstract (This volume will contain the State Tables down to the Tahsils and all cities, town-groups and towns). PART II .B--Economic Tables (B Series, Table I-IX). Likely to be in two sub-parts. PART II C.-Cultural and Migration Tables (0 and D Series). Likely to be in two sub-parts. PART III--Household Economic Table-Based on Household Schedules. (B Series. Table X to XVI). PART IV~Housing and Establishment Tables (E Series Tables) including Subsidiary Tables and Report on Housing and Establishment Tables. PART V A-Report and Tables on Scheduled Castes and Scheduled Tribes (SCT. se and ST Series Tables). PART V R-Reprints from old Census Reports and Castes and Tribes. PART VI-Village Survey Monographs. To be in several sub-parts. PART VII-Survey of Handicrafts-To be in several sub-parUo. PART VIII A-Administration Report-Enumeration. PART VIII B-Administration Report-Tabulation. PART IX-Maps. PREFACE A census administration report should be considered useful not so much for the "do's' that it may throw light upon as for the 'donts' which, in retrospect, stand out more clearly. -

Official Gazette Government Of· Goa, Daman· and Diu
:1 REGD. GOA - 51 Panaji, 4th July, 1977 (Asadha 13, 1899) SERIES I No. 13 OFFICIAL GAZETTE GOVERNMENT OF· GOA, DAMAN· AND DIU EXT ftf\O R DIN 1\ RY No. 2 Sections GOVERNMENT OP GOA. DAMAN 8. Preparation of draft statem€llt as regards vacant AND DIU land held in eXcess 'Of ceiling 11mit. 9. Final statement. law and Judiciary Department 10. AcqUisition ()f vacant land in excess of ceIling limit. 11. Payment of amount for vacant land acquired~ . 12. Constitution of Urban Land Tribunal and appeal to Notification Urban Land Tribunal. LD/1ILS6/76 13. Second appeal to High Court. The following Central Bill the Urban Land (Cei. U. Mode of payment of amount. ling and Regulation) Act, 1976 which was recently 15. Ceiling. limit on future acquisition by inheritance, passed by the Parliament and aslSented to by the bequest or by sale in execution of decrees, etc. President of India on 17·2·76 and published in the 16. Certain persons to file statements when the Act is {;azette of India Part II, Section I dated 17·2·76 adopted subsequently by any State. is hereby republished for general informatfun of the 17. Power to enter upon any ;vacant land. :public. 18: Penalty for concealment, etc., of particulans of vacant land. B. S. Subbanna, Under Secretary (Law). 19. Chapter Dot to apply to certain vacant lands. Panaji, 22nd June, 1976. 20. Power to exempt. 21. Excess vacant land not to be treated as excess in certain cases.. The Urban Land (Ceiling and Reglilation) Act. 1976 22. Retention of vacant land under certain circum.s.tailces.