Fuels and Fire Behavior Dynamics in Bark Beetle-Attacked Forests in Western North America and Implications for Fire Management
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Wildland Fire Incident Management Field Guide
A publication of the National Wildfire Coordinating Group Wildland Fire Incident Management Field Guide PMS 210 April 2013 Wildland Fire Incident Management Field Guide April 2013 PMS 210 Sponsored for NWCG publication by the NWCG Operations and Workforce Development Committee. Comments regarding the content of this product should be directed to the Operations and Workforce Development Committee, contact and other information about this committee is located on the NWCG Web site at http://www.nwcg.gov. Questions and comments may also be emailed to [email protected]. This product is available electronically from the NWCG Web site at http://www.nwcg.gov. Previous editions: this product replaces PMS 410-1, Fireline Handbook, NWCG Handbook 3, March 2004. The National Wildfire Coordinating Group (NWCG) has approved the contents of this product for the guidance of its member agencies and is not responsible for the interpretation or use of this information by anyone else. NWCG’s intent is to specifically identify all copyrighted content used in NWCG products. All other NWCG information is in the public domain. Use of public domain information, including copying, is permitted. Use of NWCG information within another document is permitted, if NWCG information is accurately credited to the NWCG. The NWCG logo may not be used except on NWCG-authorized information. “National Wildfire Coordinating Group,” “NWCG,” and the NWCG logo are trademarks of the National Wildfire Coordinating Group. The use of trade, firm, or corporation names or trademarks in this product is for the information and convenience of the reader and does not constitute an endorsement by the National Wildfire Coordinating Group or its member agencies of any product or service to the exclusion of others that may be suitable. -

Fire and Aviation Level One Report
Part 1 of 2 Missoula Fire and Aviation USDA Forest Service • Technology and Development Program • 5100 Fire, 5700 Aviation Technical Services— community through their involve- ment in groups such as the Fire and Aviation National Fire Protection Associa- Management tion (NFPA), National Wildfire Coordinating Group, and National Technical services provided by Fire Equipment Systems (NFES) MTDC’s Fire and Aviation committee. By maintaining a close Management Program are used to relationship with workers in the transfer information and technol- field—those folks in the yellow ogy from specialists to the field shirts—MTDC anticipates new units. MTDC personnel stay needs, enabling the Forest Service current on technologies and issues to provide a safer, more productive that affect the wildland firefighting work environment. Dick Mangan, Fire and Aviation Program Leader Dick has been Program Leader for Fire, Aviation, and Residues at MTDC since 1989. Before coming to the center, he spent more than National 20 years working on Ranger Districts and National Forests in Oregon and Wildfire Washington, participating in the full Coordinating range of wildland fire activities. He serves on the National Wildfire Group Coordinating Group (NWCG) Fire Equipment and Safety and Health Working Teams and is chairperson of the National Fire Protection The National Wild- Association (NFPA) 1977 Technical fire Coordinating Committee for Wildland Fire Group (NWCG) is an Personal Protective Clothing and organization whose Equipment. Dick remains active in membership includes the field, representing MTDC on fire entrapment investigations and the National Associa- serving as Operations Section Chief tion of State Foresters, on a National Type 1 Overhead Team. -

Glossary of Fire Weather Terms
2021 Arkansas Fire Weather Operating Plan National Weather Service Forecast Office Little Rock, Arkansas 1 2021 Arkansas Fire Weather Operating Plan National Weather Service Forecast Office – Little Rock 8400 Remount Road North Little Rock, AR 72118 Joseph C Goudsward - Senior Forecaster Incident Meteorologist (IMET) 2 Table of Contents Chapter Page Introduction 4 Important Changes 6 General Information 7 Role of the National Weather Service 8 Red Flag Program 10 Basic vs. Special Services 11 Forecast Products 14 Spot Forecasts 31 NFDRS Forecasts 43 Update Policy 48 NWS Offices and Responsibilities 49 Appendices #1 Wildland Fire terminology 61 #2 Southern Region Contacts and IMETs 79 #3 Arkansas Fire Weather Zones 82 #4 National Fire Plan 83 #5 Fire weather Links 88 #6 NFDRS Sites 89 #7 Contact Information 90 #8 Forecast Products 97 3 Introduction This document is the latest Arkansas Fire Weather Operating Plan. It serves to consolidate the fire weather services provided by National Weather Service (NWS) offices covering the state. The purpose of this operating plan can be broken down into three distinct areas. The first is to consolidate all the fire weather services provided by the five NWS offices covering the state of Arkansas. The second purpose is to describe the services available to all land management agencies in Arkansas. The final purpose is to provide information and guidelines to the forecasting staff at the five NWS offices to ensure that consistent information is given to their customers. Customers of NWS fire weather products and services in Arkansas must understand that the products and policies contained within may differ slightly based on local policy and procedure. -

Mid-Plains Interagency Handcrew Standard Operations Guide
MID-PLAINS INTERAGENCY HANDCREW STANDARD OPERATIONS GUIDE MID-PLAINS INTERAGENCY CREW Service, Growth, Leadership 1 Service, Growth, Leadership 2 Table of Contents Purpose ........................................................................................................................ 4 Defining “Interagency” ............................................................................................... 4 Mission Statement ....................................................................................................... 4 Code of Conduct ......................................................................................................... 4 Crew Guiding Principals ............................................................................................. 4 Safety .......................................................................................................................... 5 Briefing Checklist ....................................................................................................... 5 Maintaining Reliability and Performance ................................................................... 5 Driving / Travel........................................................................................................... 5 Qualifications .............................................................................................................. 6 Organization ................................................................................................................ 6 Schedule ..................................................................................................................... -

Training with a Practice Fire Shelter
"Zero Tolerance" memo to all employees from Secretary of Agriculture Dan Glickman and Secretary of the Interior Bruce Babbitt. " <- WASHINGTON ->:. '-".. ---.'1. S-.• THE SECRETARY OF AGRICULTURE THE SECRETARY OF THE INTERIOA TO ALLEMPLOYEES 1994 was a tragic year for wildland fire. Even more sobering is that without the judgment and commitment to safety demonstrated by fuefighting personnel throughout the Nation, our losses could have been even greater. Important lessons were learned, including an affinnation thar agency personnel at all levels. and not just those directly involved in fire suppression, must demonstrate a commitmentto safety. We are committed to "Zero Tolerance" ofcarelessness and unsafe actions. The commitment to and accountability for safety is a joint responsibility of firefighters. managers and administrators. No resource or property values are worth endangering people. All land management plans and all Dan Glickman suppression plans and actions must reflect this commitment. Individuals must be personally committed and responsible for their own performance and accountability. Please join us in adopting firefighting's code of safe practices: Safety Comes First on Every Fire, Every Time. The Ten Standard Fire Orders are Firm. We Don't Break Them; We Don't Bend Them. An Firefighters have the Right to a Safe Assignment Every Firefighter, Every Pireline Supervisor, Every Fire Manager, and Every Agency Administrator has the Responsibility to Ensure Compliance with Established Safe Firefighting Practices. ?!Z~ secretary of the Interior BruceBabbitt This issue of Fire ManagementNotes is Fire Management Notes is published by the Forest Service of the U.S. Department ofAgriculture, Washington, DC. the first of two focusing on the safety and The Secretary of Agriculture has determined that the publication of this periodical is necessary in the transaction of the public business required by law of this Department. -
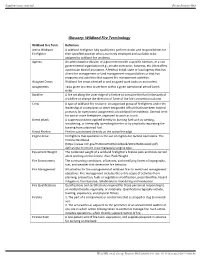
Glossary: Wildland Fire Terminology
Supplementary material Occup Environ Med Glossary: Wildland Fire Terminology Wildland Fire Term Definition Active Wildland A wildland firefighter fully qualified to perform duties and responsibilities for Firefighter their specified position who is currently employed and available to be assigned to wildland fire incidents. Agency An administrative division of a government with a specific function, or a non- governmental organization (e.g., private contractor, business, etc.) that offers a particular kind of assistance. A federal, tribal, state or local agency that has direct fire management or land management responsibilities or that has programs and activities that support fire management activities. Assigned Crews Wildland fire crews checked in and assigned work tasks on an incident. Assignments Tasks given to crews to perform within a given operational period (work shift). Backfire A fire set along the inner edge of a fireline to consume the fuel in the path of a wildfire or change the direction of force of the fire's convection column. Crew A type of wildland fire resource. An organized group of firefighters under the leadership of a crew boss or other designated official that have been trained primarily for operational assignments on wildland fire incidents. General term for two or more firefighters organized to work as a unit. Direct attack A suppression tactic applied directly to burning fuel such as wetting, smothering, or chemically quenching the fire or by physically separating the burning from unburned fuel. Direct Fireline Fireline constructed directly on the active fire edge. Engine Crew Firefighters that specialize in the use of engines for tactical operations. The Fireline Handbook (https://www.nifc.gov/PUBLICATIONS/redbook/2019/RedBookAll.pdf) defines the minimum crew makeup by engine type. -

Frequently Asked Questions About Fire Shelters 14 May 2019
National Wildfire Coordinating Group Fire Shelter Subcommittee Frequently Asked Questions about Fire Shelters 14 May 2019 GENERAL What is a fire shelter? The fire shelter is an aluminized cloth tent that offers protection in a wildfire entrapment situation by reflecting radiant heat and providing a volume of breathable air. Additional information can be found at the fire shelter website. When did firefighters begin using fire shelters? Wildland firefighters have carried fire shelters since their development in the 1960s. The current model has been in use by firefighters since 2003. Are all federal, state, and local agency firefighters required to carry a fire shelter? Wildland firefighters were first required to carry fire shelters as personal protective equipment in 1977. The NWCG member agencies have all required the current model since 2010. When should a fire shelter be used? Firefighters train extensively in fire entrapment avoidance, so fire shelters are used in the very rare occurrence of a fire entrapment where firefighters feel the shelter is needed for protection from heat, smoke, and/or ember showers. How does the fire shelter work? The fire shelter provides protection primarily by reflecting radiant heat and trapping breathable air inside. The current shelter is comprised of two layers. The outer layer is woven silica laminated to aluminum foil. The foil reflects radiant heat and the silica cloth slows the transfer of heat to the inside of the shelter. An inner layer is fiberglass laminated to aluminum foil. The inner layer of foil prevents heat from being reradiated inside the shelter, and it prevents gases from entering the shelter. -

Twisp River PPE and Equipment Analysis Report
United States Department of Agriculture USDA Forest Service Twisp River Fire Personal Protective Equipment (PPE) Analysis and Fire Equipment Report To accompany Twisp River Fire Fatalities and Entrapments Learning Review Narrative Fall 2016 Twisp River Fire Personal Protective Equipment (PPE) Analysis and Fire Equipment Report Contents Twisp River Fire Personal Protective Equipment (PPE) Analysis ................................................................... 2 Forest Service Engine 642 PPE ..................................................................................................... 2 Flame-resistant (FR) Clothing ............................................................................................................... 3 Helmet .................................................................................................................................................. 5 Boots .................................................................................................................................................... 6 FD6 Engine 4, COOP Engine 1, and Contractor Engine 1 ............................................................. 6 Dozer Group ................................................................................................................................ 7 Dozer Group FR Clothing and Personal Protective Equipment ................................................... 8 Fire shelters and PVC bags .................................................................................................................. -

Fireterminology.Pdf
Abandonment: Abandonment occurs when an emergency responder begins treatment of a patient and the leaves the patient or discontinues treatment prior to arrival of an equally or higher trained responder. Abrasion: A scrape or brush of the skin usually making it reddish in color and resulting in minor capillary bleeding. Absolute Pressure: The measurement of pressure, including atmospheric pressure. Measured in pound per square inch absolute. Absorption: A defensive method of controlling a spill by applying a material that absorbs the spilled material. Accelerant: Flammable fuel (often liquid) used by some arsonists to increase size or intensity of fire. Accelerator: A device to speed the operation of the dry sprinkler valve by detecting the decrease in air pressure resulting in acceleration of water flow to sprinkler heads. Accountability: The process of emergency responders (fire, police, emergency medical, etc...) checking in as being on-scene during an incident to an incident commander or accountability officer. Through the accountability system, each person is tracked throughout the incident until released from the scene by the incident commander or accountability officer. This is becoming a standard in the emergency services arena primarily for the safety of emergency personnel. Adapter: A device that adapts or changes one type of hose thread, type or size to another. It allows for connection of hoses and pipes of incompatible diameter, thread, or gender. May contain combinations, such as a double-female reducer. Adapters between multiple hoses are called wye, Siamese, or distributor. Administrative Warrant: An order issued by a magistrate that grants authority for fire personnel to enter private property for the purpose of conducting a fire prevention inspection or similar purpose. -

New Standards for Wildland Firefighting Protective Clothing and Equipment Dick Mangan, Program Leader
United States Department of Agriculture Fire Forest Service Technology & Development Program August 1993 Tech Tips 5100 9351 2340–MTDC New Standards for Wildland Firefighting Protective Clothing and Equipment Dick Mangan, Program Leader At its May 1993 annual meeting, the National Fire Protection Association (NFPA) approved newly developed standards on protective clothing and equipment for wildland firefighting (NFPA 1977-1993 edition). This new NFPA standard is the first to specifically address the wildland firefighter. It results from a 4-year effort to develop minimum design and performance criteria and testing methods for evaluating the thermal protection of flame-resistant clothing without inducing extraordinary heat load. Items covered by the NFPA 1977 standard include the protective helmet (hardhat), protective clothing (shirt, trousers, and one- piece coverall), footwear, gloves, and fire shelter. The subcommittee that recommended the draft standards to the NFPA Technical Committee on Fire Service Protective Clothing and Equipment represented a broad cross-section of agencies, organizations, and manufacturers interested in wildland fire safety. USDA Forest Service representation included fire personnel from both the West and Thermal protective clothing and equipment. South, as well as technical specialists from the Missoula Technology and were included for clothing items, because MTDC’s Development Center (MTDC) in Montana. State extensive study of fire entrapment shows that loose- Forestry organization from the Northeast and West, fitting clothing is oftentimes just as important as the testing laboratories, regulatory agencies, flame-resistance of materials in preventing serious labor unions, manufacturers, and suppliers also burn injuries. Wearing an oversized protective served on the subcommittee. garment of the same nominal waist size as normal workwear is the best way to ensure the loose fit that is The standards for clothing, protective helmet, boots a critical safety feature of these garments. -

Constructing Fireline (Wildland) Page 1 Developed by Engineer John Sanfilippo September 2011 • National Wildfire Coordinatio
C ONSTRUCTING F IRE L INE (WILDLAND) (1.1) Developed by Engineer John Sanfilippo September 2011 National Wildfire Coordination Group (NWCG) – Incident Response Pocket Guide (IRPG) 2010 Ed. Standard Firefighting Orders (F.O.) and Watch Out Situations (W.O.) LFR Playbook – Wildland Fire Operating Guide (December 2006) T ASK S KILL DESCRIPTION AND DETAIL Firefighter Safety F.O. #10 – Fight fire aggressively, having provided for safety first. First identify L.C.E.S. and brief all members to ensure they are known and understood (F.O.’s #4, #5, #7, and #8). Size up the fire and know what the fire is doing or get in contact with someone that can (F.O. #2). All fire line must begin with an adequate anchor point (W.O. #8). Fire Line Location Always start with an anchor point. Will the location of your anchor point keep the fire from hooking around behind you? Answer this question or make adjustments to answer it. Consider another point to begin if you can’t answer it. Will some improvements work to answer the question? Usually, it is best to begin downhill of the fire. Look for natural features to help you; utilize wetlands, rock outcroppings, roads or trails to start with. Constructing Fireline (Wildland) Page 1 Once the anchor point is established (Benchmark – Anchor Point Established), time to build line on the fire flanks. Direct attack with one foot in the green and one foot in the black is safest and preferred. Try to keep the line as straight as possible and utilize natural features to lessen the workload. -

Wildland Fire Equipment 2019 Visit Us on the Web At: Wildland Fire Equipment Products from DLA
DEFENSE LOGISTICS AGENCY Wildland Fire Equipment 2019 Visit us on the Web at: https://www.fedmall.mil/ Wildland Fire Equipment Products from DLA We are proud to present this special catalog spotlighting wildland fire equipment Contents available from FedMall. From hand tools to protective gear, FedMall offers a variety of products that carry our service guarantees: About This Catalog ........................................ 2 • Easy Requisitions – See It. Buy It. Get It • Flexible – Featuring Multiple Ordering Options • Reliable – Delivering the ProductsYou Need,WhereYou Need Them Wildland Fire Equipment • Anywhere – Assuring Global Delivery Hand Tools ................................................................................... 4 • Anytime – Supporting 24/7 Purchasing Sheaths .......................................................................................... 5 • More – Offering Extensive Product Selection Handles and Handle Wedges ..................................................... 5 • Safe – Providing Easy Compliance Hoses and Gaskets ....................................................................... 6 The items in this publication are also available online. Visit us at: Water-Handling Accessories ...................................................... 7 https://www.fedmall.mil Pumps and Accessories ............................................................ 11 Chainsaw Accessories ............................................................... 13 Fuel Storage ...............................................................................