Further Contribution to the Siwalik Flora Froul the Koilabas Area, Western Nepal
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Volume 52-1, February
VOLUME 52.1 SPRING 2012 UN NGO Consultative Status ESCO & DPI 1981 2011-2012 BOARD MESSAGE FROM THE PRESIDENT EXECUTIVE COMMITTEE President: Dr. Ludwig F. Lowenstein, England MESSAGE FROM THE PRESIDENT Fair Oak 004423 80692621 Dear Colleagues, friends and members of our family ICP, President Elect: Dr.Tara Pir, USA, Los Angeles, CA Another festival season and year is behind us and ahead of us, espe- (213) 381-1250 cially in the Northern Hemisphere are cold days and nights while in the Past President: : Dr.Ann Marie O’Roark, USA Southern Hemisphere it should be lovely and warm. Those who live in St. Augustine, FL the Southern Hemisphere are envied by those shivering in the cold. (904) 461 3382 Treasurer: Dr. Gerald L. Ga- Things are well on the way for attendance at conferences in mache, USA, St. Augustine, South Africa and Sevilla. Those who are coming to South Africa I FL would suggest the Fountain Hotel which is reasonably priced and of (904 ) 824- 5668 Secretary:Dr. Donna Goetz, good quality. It is near the convention centre. I would also consider the USA, Lombard, IL Hotel Viapol, in Sevilla which is also very convenient and near the University where the 630) 627-4969 ICP conference will be held. I will be arriving a day early and staying a day later after the DIRECTORS AT LARGE conference to meet you all and have a pre-convention informal get-together. As usual I reit- Term Expires in 2012 erate the importance of our finding new members who will hopefully be coming to the con- Prof. -

'Grand Theft Archive': a Quantitative Analysis of the State of Computer
Gooding, P. and Terras, M. (2008) "„Grand Theft Archive‟: a quantitative analysis of the current state of computer game preservation". The International Journal of Digital Curation. Issue 2, Volume 3, 2008. http://www.ijdc.net/ijdc/article/view/85/90 1 ‘Grand Theft Archive’: A Quantitative Analysis of the Current State of Computer Game Preservation Paul Gooding, Librarian, BBC Sports Library, London Melissa Terras, School of Library, Archive and Information Studies, University College London November 2008 Abstract Computer games, like other digital media, are extremely vulnerable to long-term loss, yet little work has been done to preserve them. As a result we are experiencing large-scale loss of the early years of gaming history. Computer games are an important part of modern popular culture, and yet are afforded little of the respect bestowed upon established media such as books, film, television and music. We must understand the reasons for the current lack of computer game preservation in order to devise strategies for the future. Computer game history is a difficult area to work in, because it is impossible to know what has been lost already, and early records are often incomplete. This paper uses the information that is available to analyse the current status of computer game preservation, specifically in the UK. It makes a quantitative analysis of the preservation status of computer games, and finds that games are already in a vulnerable state. It proposes that work should be done to compile accurate metadata on computer games and to analyse more closely the exact scale of data loss, while suggesting strategies to overcome the barriers that currently exist. -

DVD / CD / MP Player
Owner’s Manual Owner’s ® DVD / CD / MP Player /MP /CD DVD T517 РУССКИЙ SVENSKA NEDERLANDS DEUTSCH ITALIANO ESPAÑOL FRANÇAIS ENGLISH IMPORTANT SAFETY INSTRUCTIONS ENGLISH SAVE THESE INSTRUCTIONS FOR LATER USE. 14 Outdoor Antenna Grounding - If an outside antenna or cable system FOLLOW ALL WARNINGS AND INSTRUCTIONS MARKED ON THE is connected to the product, be sure the antenna or cable system is AUDIO EQUIPMENT. grounded so as to provide some protection against voltage surges and built-up static charges. Article 810 of the National Electrical Code, 1 Read instructions - All the safety and operating instructions should be ANSI/NFPA 70, provides information with regard to proper grounding read before the product is operated. of the mast and supporting structure, grounding of the lead-in wire 2 Retain instructions - The safety and operating instructions should be to an antenna discharge unit, size of grounding conductors, location FRANÇAIS ESPAÑOL retained for future reference. of antenna discharge unit, connection to grounding electrodes, and 3 Heed Warnings - All warnings on the product and in the operating requirements for the grounding electrode. instructions should be adhered to. 4 Follow Instructions - All operating and use instructions should be NOTE TO CATV SYSTEM INSTALLER followed. This reminder is provided to call the CATV system installer’s attention to Section 5 Cleaning - Unplug this product from the wall outlet before cleaning. 820-40 of the NEC which provides guidelines for proper grounding and, in Do not use liquid cleaners or aerosol cleaners. Use a damp cloth for particular, specifies that the cable ground shall be connected to the grounding cleaning. -

Vc-300Hd-Introduction.Pdf
Now that digital broadcasting service areas are expanding, and with the appearance of high-definition content on Blu-ray and HD DVD media, there is a strong demand for high definition content production. Until these recent developments, HD content production had been comparatively simple, with the source of input being mainly limited to HD camera recordings. With television shifting to high definition and the rapid spread of high-definition video cameras for consumer use, location shooting in consumer HDV format with inexpensive cameras is becoming more popular. Meanwhile the transition to a tapeless era for broadcasting is underway. Overall, an extremely complex situation has arisen. If this had been a complete shift towards digital data in files handled by codecs, IP and the world of networking, this may not have presented so many challenges. In reality, we cannot ignore the world of real-time transmissions in which data streams of various formats are distributed via cable connection. When viewed as, “Hi-Def”, the various HD formats probably look the same to many. In reality, however, video from different sources are likely to have different resolutions, bit rates, and frame rates. Moreover, when these are transmitted, it may be physically impossible to access files with different formats. It is getting so that you can’t easily manage simple tasks like dubbing or monitoring. What is needed for this world of many formats is a multi-format converter that can convert from many formats to any other form. While it is possible to get devices that can specifically convert between just about any of the existing formats, for example between component and SDI, they are not cost effective or bi-directional. -

Liberal Arts Science $600 Million in Support of Undergraduate Science Education
Janelia Update |||| Roger Tsien |||| Ask a Scientist SUMMER 2004 www.hhmi.org/bulletin LIBERAL ARTS SCIENCE In science and teaching— and preparing future investigators—liberal arts colleges earn an A+. C O N T E N T S Summer 2004 || Volume 17 Number 2 FEATURES 22 10 10 A Wellspring of Scientists [COVER STORY] When it comes to producing science Ph.D.s, liberal arts colleges are at the head of the class. By Christopher Connell 22 Cells Aglow Combining aesthetics with shrewd science, Roger Tsien found a bet- ter way to look at cells—and helped to revolutionize several scientif-ic disciplines. By Diana Steele 28 Night Science Like to take risks and tackle intractable problems? As construction motors on at Janelia Farm, the call is out for venturesome scientists with big research ideas. By Mary Beth Gardiner DEPARTMENTS 02 I N S T I T U T E N E W S HHMI Announces New 34 Investigator Competition | Undergraduate Science: $50 Million in New Grants 03 PRESIDENT’S LETTER The Scientific Apprenticeship U P F R O N T 04 New Discoveries Propel Stem Cell Research 06 Sleeper’s Hold on Science 08 Ask a Scientist 27 I N T E R V I E W Toward Détente on Stem Cell Research 33 G R A N T S Extending hhmi’s Global Outreach | Institute Awards Two Grants for Science Education Programs 34 INSTITUTE NEWS Bye-Bye Bio 101 NEWS & NOTES 36 Saving the Children 37 Six Antigens at a Time 38 The Emergence of Resistance 40 39 Hidden Potential 39 Remembering Santiago 40 Models and Mentors 41 Tracking the Transgenic Fly 42 Conduct Beyond Reproach 43 The 1918 Flu: Case Solved 44 HHMI LAB BOOK 46 N O T A B E N E 49 INSIDE HHMI Dollars and Sense ON THE COVER: Nancy H. -
Mega Unlock Dvd Player All Multi Region Code Zone Free
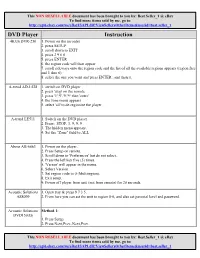
This NON RESELLABLE document has been brought to you by: Best.Seller_1 @ eBay To find more items sold by me, go to: http://cgi6.ebay.com/ws/eBayISAPI.dll?ViewSellersOtherItems&userid=best.seller_1 DVD Player Instruction 4KUS DVR-230 1. Power on the recorder 2. press SETUP 3. scroll down to EXIT 4. press 2 9 6 0 5. press ENTER 6. the region code will then appear 7. scroll sideways onto the region code and the list of all the available regions appears (region free and 1 thro 6) 8. select the one you want and press ENTER...and thats it A-trend AD-L528 1. switch on DVD player 2. press 'stop' on the remote 3. press '1','9','9','9' then 'enter' 4. the zone menu appears 5. select 'all' to de-regionize the player A-trend LE511 1. Switch on the DVD player 2. Press: STOP, 1, 9, 9, 9 3. The hidden menu appears. 4. Set the "Zone" field to ALL Aboss AB-6863 1. Power on the player. 2. Press Setup on remote. 3. Scroll down to 'Preferences' but do not select. 4. Press the left key five (5) times. 5. 'Verson' will appear in the menu. 6. Select Version. 7. Set region code to 0 (Multiregion). 8. Exit setup. 9. Power off player from unit (not from remote) for 20 seconds. Acoustic Solutions 1. Open tray & press 9 7 3 5. AS8099 2. From here you can set the unit to region 0-6, and also set parental level and password. Acoustic Solutions Method 1: DVD150AS 1. -

COVID-19 Socio-Economic Impact Assessment
COVID-19 Socio-Economic Impact Assessment COVID-19 Socio-Economic Impact Assessment Serbia Covid19 Socio Economic Impact Assessment - Sept 2020 1 FOREWORD The year 2020 will be remembered as a turning point in the This report provides a comprehensive overview of COVID- history of modern society. The year when the whole world - 19’s impact in Serbia. It captures the situation in the first six every one of us - was impacted by a crisis that has had far months following the outbreak, based on official data and reaching consequences on our lives, bringing global uncer- specific surveys conducted by UN agencies and other civil tainty and challenges not seen since World War II. sector research. The report serves to inform the choices we make and actions we take in the coming period. These deci- While the coronavirus (“COVID-19”) has brought intense sions will determine the outcome of this pandemic and the suffering to many families and communities in Serbia and development of the region for decades to come. around the world, it has also brought some unforeseen positive side effects, from clear blue skies in dense urban This report is the result of a broad, collaborative effort in- volving UN agencies and programs, the Government of Ser- environments to unprecedented levels of innovation and bia, and representatives of both businesses and civil soci- solidarity. ety. It will be used by the UN family and beyond to identify and propose short and long-term recovery solutions, with We have already learned a lot from this crisis, and we know a focus on the most vulnerable groups and those who risk that it has the potential to induce a change for the better. -

Non-Military Security and Global Order the Impact of Extremism, Violence and Chaos on National and International Security
Non-Military Security and Global Order The Impact of Extremism, Violence and Chaos on National and International Security Peter Chalk Non-Military Security and Global Order Also by Peter Chalk WEST EUROPEAN TERRORISM AND COUNTER-TERRORISM: The Evolving Dynamic Non-Military Security and Global Order The Impact of Extremism, Violence and Chaos on National and International Security Peter Chalk National Security Research Division RAND Corporation Arlington, Virginia First published in Great Britain 2000 by MACMILLAN PRESS LTD Houndmills, Basingstoke, Hampshire RG21 6XS and London Companies and representatives throughout the world A catalogue record for this book is available from the British Library. ISBN 0–333–77373–X First published in the United States of America 2000 by ST. MARTIN’S PRESS, LLC, Scholarly and Reference Division, 175 Fifth Avenue, New York, N.Y. 10010 ISBN 0–312–23167–9 Library of Congress Cataloging-in-Publication Data Chalk, Peter. Non-military security and global order : the impact of extremism, violence, and chaos on national and international security / Peter Chalk. p. cm. Includes bibliographical references and index. ISBN 0–312–23167–9 (cloth) 1. Internal security. 2. Security, International. I. Title. HV6419 .C53 2000 363.3'2—dc21 00–021169 © Peter Chalk 2000 All rights reserved. No reproduction, copy or transmission of this publication may be made without written permission. No paragraph of this publication may be reproduced, copied or transmitted save with written permission or in accordance with the provisions of the Copyright, Designs and Patents Act 1988, or under the terms of any licence permitting limited copying issued by the Copyright Licensing Agency, 90 Tottenham Court Road, London W1P 0LP. -

BOOKS, Dvds, TOOLS & PRODUCTS
FREE pass it on! 2011 BOOKS, DVDs, TOOLS & PRODUCTS 100s .. of . E BESTSELLERS G see D E inside! G IN T T U C . .. G IN G N A H C E IF . L E.. TIV PRACTICAL... INFORMA chosen by the staff of Permaculture Magazine www.green-shopping.co.uk ED BY R T E H E W GS S WELCOME TO GREEN SHOPPING 2011 O U P N his new edition of the Green Shopping catalogue (est. 1994) is full of our bestsellers. All items are chosen Tby the Permaculture Magazine team. The books we sell we’ve personally found useful, inspirational or they’ve been recommended to us. All of the tools and products we sell are personally tested by us or by relevant experts in a particular field. We believe that this level of attention is essential to ensure quality of selection, but we admit we enjoy our work! We are practical people with a passion for the subjects – it is fun finding useful, inspiring books, robust specialist tools, outdoor cooking equipment and great gifts. With our selection, we stick to some basic but essential criteria. We are foremost a green catalogue run by perma- culture people. We sell genuinely practical, useful or educational materials. Our products are made by small companies from ethically sourced materials. If we import it is because the product cannot be sourced more ‘locally’. We don’t sell goods from sweatshops. We welcome your feedback and ideas for new items and we are always at the Above: Henry, Mary and Holly test out the practicality of the Bon-Fire end of an email or a phone during office hours to offer a Outdoor Kitchen Set (see page 26). -

DVD / CD / MP3 Player ENGLISH FRANÇAIS DEUTSCH NEDERLANDS ESPAÑOL
T515_NAD_English_Final.qxd 5/7/06 11:45 AM Page 1 ® T515 DVD / CD / MP3 Player ENGLISH FRANÇAIS DEUTSCH NEDERLANDS ESPAÑOL Owner’s Manual Manuel de l'utilisateur Bedienungsanleitung ITALIANO Gebruikershandleiding Manual del Usuario Manuale delle Istruzioni PORTUGUÊS Manual do Proprietário Bruksanvisning SVENSKA T515_NAD_English_Final.qxd 5/7/06 11:45 AM Page 2 Introduction IMPORTANT SAFETY INSTRUCTIONS CAUTION CAUTION This apparatus should not be exposed to water (dripping or splashing) RISK OF ELECTRIC SHOCK and no objects filled with liquids, such as vases, should be placed on the DO NOT OPEN apparatus. ENGLISH FRANÇAIS FCC WARNING WARNING: TO REDUCE THE RISK OF ELECTRIC This equipment may generate or use radio frequency energy. Changes SHOCK, DO NOT REMOVE THE COVER OR BACK or modifications to this equipment may cause harmful interference OF THIS PRODUCT. THERE ARE NO USER- unless the modifications are expressly approved in the instruction SERVICEABLE PARTS INSIDE. REFER SERVICING manual. The user could lose the authority to operate this equipment if TO QUALIFIED SERVICE PERSONNEL. an unauthorized change or modification is made. WARNING: TO REDUCE THE RISK OF FIRE OR REGULATORY INFORMATION: FCC PART 15 ELECTRIC SHOCK, DO NOT EXPOSE THIS PRODUCT This product has been tested and found to comply with the limits for a TO DRIPPING OR SPLASHING WATER, RAIN, OR Class B digital device, pursuant to Part 15 of the FCC Rules. These limits MOISTURE. DO NOT PLACE OBJECTS FILLED WITH are designed to provide reasonable protection against harmful WATER SUCH AS VASES, ON THE PRODUCT. interference when the product is operated in a residential installation. -

Panasonic AG-AC160 HD Cameras Kits
This product is eligible for the AVCCAM 3 Year Warranty Repair Program. For details, see page 9 of vol.1. Operating Instructions Vol.2 Memory Card Camera-Recorder Model No. AG-AC160P AG-AC160EJ Volume 2 AG-AC160EN Note that Operation Instructions Vol.2 describes advanced operations of the Memory Card Camera-Recorder. For instructions on basic operations of the Memory Card Camera- Recorder, refer to Operating Instructions Vol.1 (printed documents) contained in the supplied CD-ROM. Before operating this product, please read the instructions carefully and save this manual for future use. M0811SI0 -YI ENGLISH VQT3T51A Contents Volume 1 Read this first! Power sources How to use the camera Adjusting the hand strap Please read before use Attaching the shoulder strap Operating precautions Attaching/removing the lens hood Attaching/removing the lens cap Fitting the eye cup Before use The remote control Turning the camera ON/OFF Before using the camera Setup menu basic operations Accessories Setting the calendar Optional accessories Menu Description of parts Setup menu structure Description of parts Reference Preparation Specifications Recharging the battery Volume 2 Shooting Digital zoom function .................................... 17 Shooting in manual mode ............................ 18 Using the viewfinder ...................................... 5 Switching to manual mode............................ 18 Using the viewfinder ....................................... 5 Setting focus (manual focus) ........................ 18 Using the LCD monitor -

Check-List of Birds of the World Volume Xiv
HARVARD UNIVERSITY LIBRARY OF THE Museum of Comparative Zoology ivtUS. COMP. ZOOU LIBRARY DEC 1 1 1968 HARVARD JUNlVERS.IX>!i CHECK-LIST OF BIRDS OF THE WORLD VOLUME XIV CHECK-LIST OF BIRDS OF THE WORLD A Continuation of the Work of James L. Peters Edited by RAYMOND A. PAYNTER, JR. In consultation with Ernst Mayr VOLUME XIV Parulidae Drepanididae Vireonidae Icteridae Fringillinae Carduelinae Estrildidae Viduinae By EMMET R. BLAKE, JAMES C. GREENWAY, JR. THOMAS R. HOWELL, GEORGE H. LOWERY, JR. ERNST MAYR, BURT L. MONROE, JR., RAYMOND A. PAYNTER, JR., AUSTIN L. RAND, AND MELVIN A. TRAYLOR CAMBRIDGE • MASSACHUSETTS MUSEUM OF COMPARATIVE ZOOLOGY 1968 COPYRIGHT, 1968 BY THE PRESIDENT AND FELLOWS OP HARVARD COLLEGE .on MUS. COM P. ZOOL. LIBRARY DEC 1 1 it.ob HARVARD UNIVERSITY^ THE HEFFERNAN PRESS, INC. WORCESTER, MASS. INTRODUCTION The eight taxa covered in this volume (Parulidae, Drepan- ididae, Vireonidae, Icteridae, Fringillidae, CardueHnae, Es- trildidae, and Viduinae) contain only about six percent of the living species of birds, but these are among the best known of the entire class. For many of these species there is comparatively abundant information on their natural his- tory, general biology, physiology, ethology, and even genet- ics. A comparable amount of data could not be found for any similar assemblage of birds. However, in spite of the abun- dance of taxonomically useful information, or more probably because of it, the systematics of these birds are among the most unsettled and controversial in the Class Aves. One would hope that each volume of the Check-list would have a useful life span of at least a few decades.