A Biogeographic Delineation of the European Alpine System Based on a Cluster
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Značajke I Evolucija 5S Rdna U Vrstama Iz Roda Pulsatilla (Mill.)
Sveučilište u Zagrebu Prirodoslovno-matematički fakultet Ivan Bakšić i Ana Dudaš Značajke i evolucija 5S rDNA u vrstama iz roda Pulsatilla (Mill.) Zagreb, 2019. Ovaj rad izrađen je na Zavodu za molekularnu biologiju Prirodoslovno-matematičkog fakulteta pod vodstvom prof. dr. sc. Višnje Besendorfer i predan je na natječaj za dodjelu Rektorove nagrade u akademskoj godini 2018./2019. Sadržaj 1. UVOD ..................................................................................................................................................1 1.1. Važnost ribosomske DNA (rDNA) u filogeniji .............................................................................1 1.2. Modeli evolucije multigenskih porodica .......................................................................................1 1.3. Značajke roda Pulsatilla ................................................................................................................2 2. OPĆI I SPECIFIČNI CILJEVI RADA ............................................................................................6 3. MATERIJALI I METODE ...............................................................................................................7 3.1. Materijal ........................................................................................................................................7 3.1.1. Biljni materijal ........................................................................................................................7 3.2. Metode ...........................................................................................................................................8 -

Relationships Among Levels of Biodiversity and the Relevance of Intraspecific Diversity in Conservation – a Project Synopsis F
ARTICLE IN PRESS Perspectives in Plant Ecology, Evolution and Systematics Perspectives in Plant Ecology, Evolution and Systematics 10 (2008) 259–281 www.elsevier.de/ppees Relationships among levels of biodiversity and the relevance of intraspecific diversity in conservation – a project synopsis F. Gugerlia,Ã, T. Englischb, H. Niklfeldb, A. Tribschc,1, Z. Mirekd, M. Ronikierd, N.E. Zimmermanna, R. Holdereggera, P. Taberlete, IntraBioDiv Consortium2,3 aWSL Swiss Federal Research Institute, Zu¨rcherstrasse 111, 8903 Birmensdorf, Switzerland bDepartment of Biogeography, University of Vienna, Rennweg 14, 1030 Wien, Austria cDepartment of Systematic and Evolutionary Botany, Rennweg 14, 1030 Wien, Austria dDepartment of Vascular Plant Systematics, Institute of Botany, Polish Academy of Science, Krako´w, Lubicz 46, 31-512 Krako´w, Poland eLaboratoire d’Ecologie Alpine (LECA), CNRS UMR 5553, University Joseph Fourier, BP 53, 2233 Rue de la Piscine, 38041 Grenoble Cedex 9, France Received 11 June 2007; received in revised form 4 June 2008; accepted 9 July 2008 Abstract The importance of the conservation of all three fundamental levels of biodiversity (ecosystems, species and genes) has been widely acknowledged, but only in recent years it has become technically feasible to consider intraspecific diversity, i.e. the genetic component to biodiversity. In order to facilitate the assessment of biodiversity, considerable efforts have been made towards identifying surrogates because the efficient evaluation of regional biodiversity would help in designating important areas for nature conservation at larger spatial scales. However, we know little about the fundamental relationships among the three levels of biodiversity, which impedes the formulation of a general, widely applicable concept of biodiversity conservation through surrogates. -

Native Or Suitable Plants City of Mccall
Native or Suitable Plants City of McCall The following list of plants is presented to assist the developer, business owner, or homeowner in selecting plants for landscaping. The list is by no means complete, but is a recommended selection of plants which are either native or have been successfully introduced to our area. Successful landscaping, however, requires much more than just the selection of plants. Unless you have some experience, it is suggested than you employ the services of a trained or otherwise experienced landscaper, arborist, or forester. For best results it is recommended that careful consideration be made in purchasing the plants from the local nurseries (i.e. Cascade, McCall, and New Meadows). Plants brought in from the Treasure Valley may not survive our local weather conditions, microsites, and higher elevations. Timing can also be a serious consideration as the plants may have already broken dormancy and can be damaged by our late frosts. Appendix B SELECTED IDAHO NATIVE PLANTS SUITABLE FOR VALLEY COUNTY GROWING CONDITIONS Trees & Shrubs Acer circinatum (Vine Maple). Shrub or small tree 15-20' tall, Pacific Northwest native. Bright scarlet-orange fall foliage. Excellent ornamental. Alnus incana (Mountain Alder). A large shrub, useful for mid to high elevation riparian plantings. Good plant for stream bank shelter and stabilization. Nitrogen fixing root system. Alnus sinuata (Sitka Alder). A shrub, 6-1 5' tall. Grows well on moist slopes or stream banks. Excellent shrub for erosion control and riparian restoration. Nitrogen fixing root system. Amelanchier alnifolia (Serviceberry). One of the earlier shrubs to blossom out in the spring. -
Dwarf Thistle, Cirsi
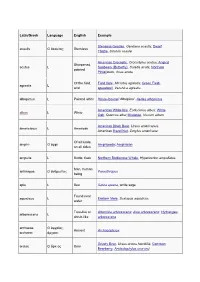
Latin/Greek Language English Example Stemless Gentian, Gentiana acaulis; Dwarf acaulis G ἄκαυλος Stemless Thistle, Cirsium acaule American Crocodile, Crocodylus acutus; Angled Sharpened, acutus L Sunbeam (Butterfly), Curetis acuta; Northern pointed Pintailduck, Anas acuta Of the field, Field Vole, Microtus agrestis; Green Field- agrestis L wild speedwell, Veronica agrestis albopictus L Painted white Hosta fortunei 'Albopicta', Aedes albopictus American White Ibis, Eudocimus albus; White albus L White Oak, Quercus alba; Mistletoe, Viscum album American Black Bear, Ursus americanus; americanus L American American Hazel Nut, Corylus americana Of all kinds, amphi- G ἀμφί Amphipoda; Amphibian on all sides ampulla L Bottle, flask Northern Bottlenose Whale, Hyperoodon ampullatus Man, human anthropos G ἄνθρωπος Paranthropus being apis L Bee Salvia apiana, white sage Found near aquaticus L Eastern Mole, Scalopus aquaticus water Tree-like or Artemisia arborescens; Aloe arborescens; Hydrangea arborescens L shrub-like arborescens archaeos, G ἀρχαῖος, Ancient Archaeopteryx archaeo- ἀρχαιο- Grizzly Bear, Ursus arctos horribilis; Common arctos G ἄρκτος Bear Bearberry, Arctostaphylos uva-ursi argentatus L Silvery Herring Gull, Larus argentatus arthron G ἄρθρον Joint Arthropoda arvensis L In the field Skylark, Alauda arvensis astron, astro-, G ἄστρον, Star Starfish (class), Asteroidea astero- ἀστρο-, ἀστερο- Acer palmatum 'Atropurpureum'; Berberis atropurpureum L Deep purple thunbergii f. atropurpurea Daphne odora 'Aureomarginata'; Taxus aureomarginata -

The Rock Garden 136 the Ro
January 2016 January 2016 THE ROCK GARDEN 136 THE ROCK GARDEN 136 January 2016 THE ROCK GARDEN Volume XXXIV Part 3 - 136 January 2016 THE ROCK GARDEN Volume XXXIV Part 3 - 136 PostalPostal Subscriptions Subscriptions from from 1st October, 1st October, 2015 2015 Postal subscriptionsPostal subscriptions are payable are payable annually annually by October by October and provide and provide membership membership of the of the SRGC untilSRGC 30 thuntil September 30th September of the following of the following year. year. SubscriptionSubscription Rates Rates UK UK OverseasOverseas Single annualSingle annual £18 £18 £23 £23 Junior Junior £3 £3 £7 £7 (under 18(under on 1 18st Oct) on 1st Oct) Family Family £21 £21 £25 £25 (Two adults(Two andadults up and to two up childrento two children under 18 under on 1 18st Oct) on 1st Oct) Three yearThree subscriptions year subscriptions are available are available at three at times three the times above the aboveannual annualrates. Renewals rates. Renewals for threefor year three subscriptions year subscriptions may only may be only made be atmade the end at the of endthe three of the year three period. year period. All subscriptionAll subscription payments payments to the club to the must club be must made be inmade GB Pounds in GB Pounds Sterling. Sterling. ChequesCheques should shouldbe made be payablemade payable to ‘The Scottishto ‘The Scottish Rock Garden Rock Garden Club’ and Club’ must and be must be drawn ondrawn a UK on bank. a UK bank. SubscriptionSubscription payments payments may be may made be throughmade through the post the by post Visa byor MastercardVisa or Mastercard providingproviding the following the following information information is sent: is sent: The longThe number long number on the cardon the card The nameThe ofname the cardholder of the cardholder as shown as onshown the cardon the card The cardThe expiry card date expiry date The cv2The 3 digit cv2 number3 digit number (from back (from of back the card) of the card) The cardholder’sThe cardholder’s signature. -

Long-Term Changes in the Composition, Ecology, and Structure of Pinus Mugo Scrubs in the Apennines (Italy)
diversity Article Long-Term Changes in the Composition, Ecology, and Structure of Pinus mugo Scrubs in the Apennines (Italy) Valentina Calabrese 1, Maria Laura Carranza 1,* ID , Alberto Evangelista 1, Marco Marchetti 2 ID , Adriano Stinca 3 ID and Angela Stanisci 1 ID 1 Envix-Lab, Dipartimento di Bioscienze e Territorio, Università degli Studi del Molise, Contrada Fonte Lappone, 86090 Pesche, Italy; [email protected] (V.C.); [email protected] (A.E.); [email protected] (A.S.) 2 Centro ArIA, Università degli Studi del Molise Via F. De Sanctis, 86100 Campobasso, Italy; [email protected] 3 Dipartimento di Scienze e Tecnologie Ambientali, Biologiche e Farmaceutiche, Università della Campania Luigi Vanvitelli, via Vivaldi 43, 81100 Caserta, Italy; [email protected] * Correspondence: [email protected]; Tel.: +39-320-479-3882 Received: 4 June 2018; Accepted: 17 July 2018; Published: 20 July 2018 Abstract: Mountain ecosystems are vulnerable because of land-use and climate change. In this study, we performed a re-visitation study using historical and newly collected vegetation plots to explore the primary trends in the floristic, ecological, and structural features of Mediterranean Pinus mugo krummholz over past decades. The plant community composition over time (1992 vs. 2016–17) was analyzed by a detrended correspondence analysis followed by a statistical comparison of time steps and an analysis of the contribution of each species to temporal differences. Ecological and structural changes were analyzed by a permutational multivariate analysis of variance followed by a post hoc comparison. We observed relevant changes in the floristic composition, structure, and ecological characteristics of Pinus mugo scrub. -

North American Rock Garden Society |
Bulletin of the American Rock Garden Society Volume 50 Number 4 Fall 1992 Cover: Gentiana paradoxa by Rob Proctor of Denver, Colorado Bulletin of the American Rock Garden Society Volume 50 Number 4 Fall 1992 Features Sorting out the Gentians, by Geoffrey Charlesworth 243 Fritillaries of Central Asia, by Josef Slegl 253 Trillium Rescue, by Don L. Jacobs 261 The Story of Fritillaria 'Martha Roderick', by W.H. de Goede 264 New Home for Rock Plants, by Elisabeth Sheldon 265 Eriogonums: Secret of the Dry Garden, by Irma Gourley 271 Preserving Rock Garden Specimens, by Karen Matthews 275 Spontaneity on the Rocks, by Panayoti Kelaidis 285 The Arctic Harebell, by J.S. DeSanto 291 Hunting for Red Helleborus niger, by Will McLewin 295 Departments Plant Portrait: Gentiana paradoxa 276 Awards 299 Books 305 Gentiana algida 242 Bulletin of the American Rock Garden Society Vol. 50(4) Sorting out the Gentians by Geoffrey Charlesworth 1 here are some genera in which tors. It is one of the hallmarks of a many of the species are considered good grower if a large patch can be good alpine plants. Androsace is such produced and maintained year after a genus, and we tend to dismiss the year, but the despair of most of us, who species that are not up to the highest have only occasionally seen a few small standard as not worth growing—for plants in our own gardens and then not instance, A. loctiflora or A. albana. It always with the astonishing color we is a mistake to make such odious associate with the species. -

Phylogeny and Biogeography of Isophyllous Species of Campanula (Campanulaceae) in the Mediterranean Area
Systematic Botany (2006), 31(4): pp. 862–880 # Copyright 2006 by the American Society of Plant Taxonomists Phylogeny and Biogeography of Isophyllous Species of Campanula (Campanulaceae) in the Mediterranean Area JEONG-MI PARK,1 SANJA KOVACˇ IC´ ,2 ZLATKO LIBER,2 WILLIAM M. M. EDDIE,3 and GERALD M. SCHNEEWEISS4,5 1Department of Systematic and Evolutionary Botany, Institute of Botany, University of Vienna, Rennweg 14, A-1030 Vienna, Austria; 2Botanical Department and Botanical Garden of the Faculty of Science, University of Zagreb, HR-10000 Zagreb, Croatia; 3Office of Lifelong Learning, University of Edinburgh, 11 Buccleuch Place, Edinburgh, EH8 9LW, Scotland, U.K.; 4Department of Biogeography and Botanical Garden, Institute of Botany, University of Vienna, Rennweg 14, A-1030 Vienna, Austria 5Author for correspondence ([email protected]) Communicating Editor: Thomas G. Lammers ABSTRACT. Sequence data from the nuclear internal transcribed spacer (ITS) were used to infer phylogenetic relationships within a morphologically, karyologically, and geographically well-defined group of species of Campanula (Campanulaceae), the Isophylla group. Although belonging to the same clade within the highly paraphyletic Campanula, the Rapunculus clade, members of the Isophylla group do not form a monophyletic group but fall into three separate clades: (i) C. elatines and C. elatinoides in the Alps; (ii) C. fragilis s.l. and C. isophylla with an amphi-Tyrrhenian distribution; and (iii) the garganica clade with an amphi-Adriatic distribution, comprised of C. fenestrellata s.l., C. garganica s.l., C. portenschlagiana, C. poscharskyana, and C. reatina. Taxa currently classified as subspecies of C. garganica (garganica, cephallenica, acarnanica) and C. fenestrellata subsp. -

Plant Conservation Report 2020
Secretariat of the CBD Technical Series No. 95 Convention on Biological Diversity 4 PLANT CONSERVATION95 REPORT 2020: A review of progress towards the Global Strategy for Plant Conservation 2011-2020 CBD Technical Series No. 95 PLANT CONSERVATION REPORT 2020: A review of progress towards the Global Strategy for Plant Conservation 2011-2020 A contribution to the fifth edition of the Global Biodiversity Outlook (GBO-5). The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the copyright holders concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. This publication may be reproduced for educational or non-profit purposes without special permission, provided acknowledgement of the source is made. The Secretariat of the Convention and Botanic Gardens Conservation International would appreciate receiving a copy of any publications that use this document as a source. Reuse of the figures is subject to permission from the original rights holders. Published by the Secretariat of the Convention on Biological Diversity in collaboration with Botanic Gardens Conservation International. ISBN 9789292257040 (print version); ISBN 9789292257057 (web version) Copyright © 2020, Secretariat of the Convention on Biological Diversity Citation: Sharrock, S. (2020). Plant Conservation Report 2020: A review of progress in implementation of the Global Strategy for Plant Conservation 2011-2020. Secretariat of the Convention on Biological Diversity, Montréal, Canada and Botanic Gardens Conservation International, Richmond, UK. Technical Series No. 95: 68 pages. For further information, contact: Secretariat of the Convention on Biological Diversity World Trade Centre, 413 Rue St. -

Development of Distribution Maps of Grassland Habitats of EUNIS Habitat Classification
Development of distribution maps of grassland habitats of EUNIS habitat classification Joop H.J. Schaminée Milan Chytrý Jürgen Dengler Stephan M. Hennekens John A.M. Janssen Borja Jiménez-Alfaro Ilona Knollová Flavia Landucci Corrado Marcenò John S. Rodwell Lubomír Tichý and data-providers Report EEA/NSS/16/005 1 Alterra, Institute within the legal entity Stichting Dienst Landbouwkundig Onderzoek Professor Joop Schaminée Stephan Hennekens Partners Professor John Rodwell, Ecologist, Lancaster, UK Professor Milan Chytrý, Masaryk University, Brno, Czech Republic Doctor Ilona Knollová, Masaryk University, Brno, Czech Republic Doctor Lubomír Tichý, Masaryk University, Brno, Czech Republic Date: 07 December 2016 Alterra Postbus 47 6700 AA Wageningen (NL) Telephone: 0317 – 48 07 00 Fax: 0317 – 41 90 00 In 2003 Alterra has implemented a certified quality management system, according to the standard ISO 9001:2008. Since 2006 Alterra works with a certified environmental care system according to the standard ISO 14001:2004. © 2014 Stichting Dienst Landbouwkundig Onderzoek All rights reserved. No part of this document may be reproduced, stored in a retrieval system, or transmitted in any form or by any means - electronic, mechanical, photocopying, recording, or otherwise - without the prior permission in writing of Stichting Dienst Landbouwkundig Onderzoek. 2 TABLE OF CONTENTS 1 Introduction 2 Scope of the project 2.1 Background 2.2 Review of the EUNIS grassland habitat types 3 Indicator species of the revised EUNIS grassland habitat types 3.1 Background -

The Gentianaceae - Volume 2: Biotechnology and Applications the Gentianaceae - Volume 2: Biotechnology and Applications Gentiana Tibetica King
Jan J. Rybczyński · Michael R. Davey Anna Mikuła Editors The Gentianaceae - Volume 2: Biotechnology and Applications The Gentianaceae - Volume 2: Biotechnology and Applications Gentiana tibetica King. (Photograph A. Mikuła) Jan J. Rybczyński • Michael R. Davey Anna Mikuła Editors The Gentianaceae - Volume 2: Biotechnology and Applications 123 Editors Jan J. Rybczyński Anna Mikuła Botanical Garden-Center for Biological Botanical Garden-Center for Biological Diversity Conservation Diversity Conservation Polish Academy of Sciences Polish Academy of Sciences Warsaw Warsaw Poland Poland Michael R. Davey Plant and Crop Sciences Division, School of Biosciences University of Nottingham Loughborough UK ISBN 978-3-642-54101-8 ISBN 978-3-642-54102-5 (eBook) DOI 10.1007/978-3-642-54102-5 Library of Congress Control Number: 2014931384 Springer Heidelberg New York Dordrecht London © Springer-Verlag Berlin Heidelberg 2015 This work is subject to copyright. All rights are reserved by the Publisher, whether the whole or part of the material is concerned, specifically the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfilms or in any other physical way, and transmission or information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology now known or hereafter developed. The use of general descriptive names, registered names, trademarks, service marks, etc. in this publication does not imply, even in the absence of a specific statement, that such names are exempt from the relevant protective laws and regulations and therefore free for general use. The publisher, the authors and the editors are safe to assume that the advice and information in this book are believed to be true and accurate at the date of publication. -

2Nd International Congress of Alpine and Arctic Botanical Gardens
Proceedings of the 2nd International Congress of Alpine and Arctic Botanical Gardens München 22-25 April 2009 CONTENTS • Introduction........................................................ 5 • Christine Freitag (Freising, Germany) Educative tools to connect an alpine garden Diversification of Collections to the surrounding vegetation......................... 35 • Katie Price (Kew, United Kingdom) • Jenny Wainwright-Klein (München, Germany) Kew’s Alpine House - what’s the point?......... 39 Experiences with the introduction of southern hemisphere alpines.............................................. 6 Research and Conservation Activities • Richard Hurstel, Pascal Salze, Christophe Per- rier, Rolland Douzet & Serge Aubert (Grenoble, • Gunter Karste (Wernigerode, Germany) France) Investigation on renaturation of the subalpine Experiences with the introduction of southern meadow vegetation on top of Brocken mountain hemisphere alpines: Southern Andes and Pata- ............................................................................. 44 gonia...................................................................... 9 • Andreas Gröger & Annette Menzel (München & • Anne Humburg (Seligenstadt, Germany) Freising, Germany) Betty Ford Alpine Gardens: the many faces of Detection of climate change impacts in alpine North America’s highest botanical garden...... 13 and arctic botanic gardens: a long-term pheno- logy observation program............................... 47 Horticultural Practices • George Nakhutsrishvili, Sh. Sikharulidze (Tbilisi, Georgia)