Nental Molluscs of Italy
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Miocene Continental Gastropods from the Southern Margin of the Swabian Alb (Baden-Württemberg, SW Germany)
N. Jb. Geol. Paläont. Abh. 287/1 (2018), 17–44 Article E Stuttgart, January 2018 Miocene continental gastropods from the southern margin of the Swabian Alb (Baden-Württemberg, SW Germany) Olaf Höltke, Rodrigo B. Salvador, and Michael W. Rasser With 6 figures Abstract: The Middle Miocene silvana-beds outcropping at the southern margin of the Swabian Alb bear a wide array of terrestrial and freshwater gastropods. Especially in the surroundings of the Emerberg and Tautschbuch hills, there are records of several collection sites. In the present work, we investigated material from the historical localities from these two regions, housed in museum collec- tions, in order to provide an updated check list, synthesizing the knowledge of the region in the hopes to spur renewed interest. In total, we report 14 freshwater and 50 land snail species from the area, but not all species reported in the literature could be confirmed. Key words: Baden-Württemberg, European Land Mammal Zone MN 5, Langhian, silvana- beds, Upper Freshwater Molasse. 1. Introduction (1923-1930). Therefore, in the absence of more driven excavations for the moment, here we investigated ma- The southern margin of the Swabian Alb, especially terial from several historical localities from these two the Emerberg and Tautschbuch hills, bears a consider- regions, housed in museum collections. Our goal is to able number of continental Miocene deposits (Fig. 1). provide an updated check list of species and localities, These are built up by the silvana-beds, a lithostrati- thus synthesizing the knowledge of the region in the graphic unit of the Miocene Upper Freshwater Molasse hopes to spur renewed interest and collection efforts. -

Spatial Predictive Distribution Modelling of Madeira's Endemic
DEPARTAMENTO DE ZOOLOGIA FACULDADE DE CIÊNCIAS E TECNOLOGIA UNIVERSIDADE DE COIMBRA Spatial predictive distribution modelling of Madeira’s endemic land snail species Dinarte Nuno Freitas Teixeira 2009 REGIÃO AUTÓNOMA DA REPÚBLICA PORTUGUESA UNIÃO EUROPEIA MADEIRA FSE DEPARTAMENTO DE ZOOLOGIA FACULDADE DE CIÊNCIAS E TECNOLOGIA UNIVERSIDADE DE COIMBRA Spatial predictive distribution modelling of Madeira’s endemic land snail species Dissertação apresentada à Universidade de Coimbra para cumprimento dos requisitos necessários à obtenção do grau de Mestre em Ecologia, realizada sob a orientação científica do Professor Doutor José Paulo Sousa (Universidade de Coimbra) e do Professor Doutor José Manuel Jesus (Universidade da Madeira). Dinarte Nuno Freitas Teixeir a 2009 O presente trabalho foi financiado pelo Centro de Ciência e Tecnologia da Madeira (CITMA), através da bolsa de Mestrado FSE BM I/2008 – 531, ao abrigo do Programa Operacional de Valorização do Potencial Humano e Coesão Social da RAM (RUMOS). REGIÃO AUTÓNOMA DA MADEIRA REPÚBLICA PORTUGUESA UNIÃO EUROPEIA FSE À Susana AGRADECIMENTOS Esta tese é o resultado de um trabalho conjunto para o qual muitos contribuíram e aos quais desejo reconhecer e deixar o meu agradecimento. Ao professor Doutor José Paulo Sousa, meu orientador, pela indispensável ajuda, paciência e orientação científica. Ao professor Doutor José Manuel Jesus, meu orientador, pela amizade e apoio desde os primeiros momentos. Pelo seu empenho, conselhos transmitidos, chamadas à razão e orientação científica o meu muito obrigado. Ao Doutor Pedro Cardoso, meu orientador e a quem muito devo, pelo constante acompanhamento e disponibilidade, amizade e orientação científica. Por tudo o que me ensinou, pela motivação e animo que sempre me transmitiu, e, acima de tudo, pela manutenção da objectividade do trabalho. -
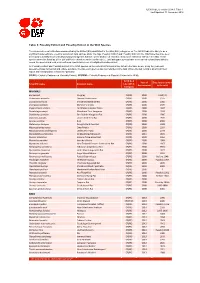
Table 9: Possibly Extinct and Possibly Extinct in the Wild Species
IUCN Red List version 2014.3: Table 9 Last Updated: 13 November 2014 Table 9: Possibly Extinct and Possibly Extinct in the Wild Species The number of recent extinctions documented by the Extinct (EX) and Extinct in the Wild (EW) categories on The IUCN Red List is likely to be a significant underestimate, even for well-known taxa such as birds. The tags 'Possibly Extinct' and 'Possibly Extinct in the Wild' have therefore been developed to identify those Critically Endangered species that are, on the balance of evidence, likely to be extinct (or extinct in the wild). These species cannot be listed as EX or EW until their extinction can be confirmed (i.e., until adequate surveys have been carried out and have failed to record the species and local or unconfirmed reports have been investigated and discounted). All 'Possibly Extinct' and 'Possibly Extinct in the Wild' species on the current IUCN Red List are listed in the table below, along the year each assessment was carried out and, where available, the date each species was last recorded in the wild. Where the last record is an unconfirmed report, last recorded date is noted as "possibly". CR(PE) - Critically Endangered (Possibly Extinct), CR(PEW) - Critically Endangered (Possibly Extinct in the Wild), IUCN Red Year of Date last recorded Scientific name Common name List (2014) Assessment in the wild Category MAMMALS Bos sauveli Kouprey CR(PE) 2008 1969/70 Crateromys australis Dinagat Crateromys CR(PE) 2008 1975 Crocidura trichura Christmas Island Shrew CR(PE) 2008 1985 Crocidura wimmeri -
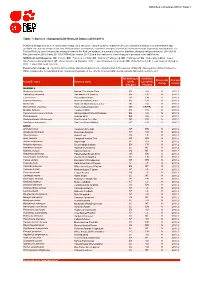
Table 7: Species Changing IUCN Red List Status (2010-2011)
IUCN Red List version 2011.2: Table 7 Table 7: Species changing IUCN Red List Status (2010-2011) Published listings of a species' status may change for a variety of reasons (genuine improvement or deterioration in status; new information being available that was not known at the time of the previous assessment; taxonomic changes; corrections to mistakes made in previous assessments, etc. To help Red List users interpret the changes between the Red List updates, a summary of species that have changed category between 2010 (IUCN Red List version 2010.4) and 2011 (IUCN Red List version 2011.2) and the reasons for these changes is provided in the table below. IUCN Red List Categories: EX - Extinct, EW - Extinct in the Wild, CR - Critically Endangered, EN - Endangered, VU - Vulnerable, LR/cd - Lower Risk/conservation dependent, NT - Near Threatened (includes LR/nt - Lower Risk/near threatened), DD - Data Deficient, LC - Least Concern (includes LR/lc - Lower Risk, least concern). Reasons for change: G - Genuine status change (genuine improvement or deterioration in the species' status); N - Non-genuine status change (i.e., status changes due to new information, improved knowledge of the criteria, incorrect data used previously, taxonomic revision, etc.) IUCN Red List IUCN Red Reason for Red List Scientific name Common name (2010) List (2011) change version Category Category MAMMALS Bradypus torquatus Maned Three-toed Sloth EN VU N 2011.1 Callicebus oenanthe San Martin Titi Monkey EN CR N 2011.1 Equus ferus Przewalski's Horse CR EN G 2011.2 -

Fauna of New Zealand Website Copy 2010, Fnz.Landcareresearch.Co.Nz
aua o ew eaa Ko te Aiaga eeke o Aoeaoa IEEAE SYSEMAICS AISOY GOU EESEAIES O ACAE ESEAC ema acae eseac ico Agicuue & Sciece Cee P O o 9 ico ew eaa K Cosy a M-C aiièe acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa EESEAIE O UIESIIES M Emeso eame o Eomoogy & Aima Ecoogy PO o ico Uiesiy ew eaa EESEAIE O MUSEUMS M ama aua Eiome eame Museum o ew eaa e aa ogaewa O o 7 Weigo ew eaa EESEAIE O OESEAS ISIUIOS awece CSIO iisio o Eomoogy GO o 17 Caea Ciy AC 1 Ausaia SEIES EIO AUA O EW EAA M C ua (ecease ue 199 acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa Fauna of New Zealand Ko te Aitanga Pepeke o Aotearoa Number / Nama 38 Naturalised terrestrial Stylommatophora (Mousca Gasooa Gay M ake acae eseac iae ag 317 amio ew eaa 4 Maaaki Whenua Ρ Ε S S ico Caeuy ew eaa 1999 Coyig © acae eseac ew eaa 1999 o a o is wok coee y coyig may e eouce o coie i ay om o y ay meas (gaic eecoic o mecaica icuig oocoyig ecoig aig iomaio eiea sysems o oewise wiou e wie emissio o e uise Caaoguig i uicaio AKE G Μ (Gay Micae 195— auase eesia Syommaooa (Mousca Gasooa / G Μ ake — ico Caeuy Maaaki Weua ess 1999 (aua o ew eaa ISS 111-533 ; o 3 IS -7-93-5 I ie 11 Seies UC 593(931 eae o uIicaio y e seies eio (a comee y eo Cosy usig comue-ase e ocessig ayou scaig a iig a acae eseac M Ae eseac Cee iae ag 917 Aucka ew eaa Māoi summay e y aco uaau Cosuas Weigo uise y Maaaki Weua ess acae eseac O o ico Caeuy Wesie //wwwmwessco/ ie y G i Weigo o coe eoceas eicuaum (ue a eigo oaa (owe (IIusao G M ake oucio o e coou Iaes was ue y e ew eaIa oey oa ue oeies eseac -

Guidelines for the Capture and Management of Digital Zoological Names Information Francisco W
Guidelines for the Capture and Management of Digital Zoological Names Information Francisco W. Welter-Schultes Version 1.1 March 2013 Suggested citation: Welter-Schultes, F.W. (2012). Guidelines for the capture and management of digital zoological names information. Version 1.1 released on March 2013. Copenhagen: Global Biodiversity Information Facility, 126 pp, ISBN: 87-92020-44-5, accessible online at http://www.gbif.org/orc/?doc_id=2784. ISBN: 87-92020-44-5 (10 digits), 978-87-92020-44-4 (13 digits). Persistent URI: http://www.gbif.org/orc/?doc_id=2784. Language: English. Copyright © F. W. Welter-Schultes & Global Biodiversity Information Facility, 2012. Disclaimer: The information, ideas, and opinions presented in this publication are those of the author and do not represent those of GBIF. License: This document is licensed under Creative Commons Attribution 3.0. Document Control: Version Description Date of release Author(s) 0.1 First complete draft. January 2012 F. W. Welter- Schultes 0.2 Document re-structured to improve February 2012 F. W. Welter- usability. Available for public Schultes & A. review. González-Talaván 1.0 First public version of the June 2012 F. W. Welter- document. Schultes 1.1 Minor editions March 2013 F. W. Welter- Schultes Cover Credit: GBIF Secretariat, 2012. Image by Levi Szekeres (Romania), obtained by stock.xchng (http://www.sxc.hu/photo/1389360). March 2013 ii Guidelines for the management of digital zoological names information Version 1.1 Table of Contents How to use this book ......................................................................... 1 SECTION I 1. Introduction ................................................................................ 2 1.1. Identifiers and the role of Linnean names ......................................... 2 1.1.1 Identifiers .................................................................................. -

Leiostyla L Ove, 1852 Mollusca, Pulmonata Новые Ископаемые Для
УДК 564.38:551.782(427+497.11) В. А. Присяжнюк Leiostyla L o v e, 1852 Mollusca, Pulmonata новые ископаемые для неогена Сербии и Украины (Представлено академиком НАН Украины П. Ф. Гожиком) Представители рода Leiostyla Love впервые встречены в сарматских отложениях Сер- бии (в известном местонахождении млекопитающих Врачевич) и в среднем сармате Украины (Волыно-Подолия, Летичевщина). Описаны виды: Leiostyla krstichae из Сербии и Leiostyla ex gr. gottschicki из Украины. В неогене Европы известно лишь несколько видов рода Leiostyla L o v e, результаты изуче- ния которых изложены в работе Дж. Манганелли и др. [1]. Это миоценовые L. gottschicki We n z, описанные В. Венцем из сармата Штейнгейма [2] и Г. Шюттом из риссоевых слоев Австрии [3] и L. austriaca We n z [4] из паннона Австрии; и плиоценовые L. priscilla Pala- dilhe [1], L. capellini S a c c o [1] и L. cf. gottschicki We n z [1]. В последнее время J. K´okay [5] определил молодой экземпляр из сармата Венгрии как L. gottschicki. Этот вид встречен нами в низах нижнего сармата Польши [6]. На мой взгляд, все перечисленные виды, кроме L. austriaca, принадлежат одной группе L. gottschicki We n z. Раковины указанного рода собраны в Центральной Сербии в озерных мергелях и глинах (аналог низов нижнего сармата) в известном местонахождении млекопитающих (Врачевич) и на Подолии (Украина, Летичевщина) в скважине у с. Чапля, также в озерных мергелях средней части среднего сармата. Материал пока хранится в Институте геологических наук НАН Украины в моей коллекции. СЕМЕЙСТВО ORCULIDAE STEENBERG, 1925 Род Leiostyla L o v e, 1852 Leiostyla ex gr. gottschicki We n z, 1921 Табл. -

Format Mitteilungen
9 Mitt. dtsch. malakozool. Ges. 86 9 – 12 Frankfurt a. M., Dezember 2011 Under Threat: The Stability of Authorships of Taxonomic Names in Malacology RUUD A. BANK Abstract: Nomenclature must be constructed in accordance with agreed rules. The International Commission on Zoological Nomenclature was founded in Leiden in September 1895. It not only produced a Code of nomencla- ture, that was refined over the years, but also provided arbitration and advice service, all with the aim of ensur- ing that every animal has one unique and universally accepted name. Name changes reduce the efficiency of biological nomenclature as a reference system. The Code was established to precisely specify the circumstances under which a name must be changed, and in what way. Name changes are only permitted if it is necessitated by a correction of nomenclatural error, by a change in classification, or by a correction of a past misidentification. Also authorships are regulated by the Code, mainly by Article 50. In a recent paper by WELTER-SCHULTES this Article is interpreted in a way that is different from previous interpretations by the zoological (malacological) community, leading to major changes in authorships. It is here argued that his alternative interpretations (1) are not in line with the spirit of the Code, and (2) will not serve the stability of nomenclature. It is important that interpretation and application of the existing rules be objective, consistent, and clear. Keywords: authorships, malacology, nomenclature, Code, ICZN, Article 50, Pisidium Zusammenfassung: In der Nomenklatur müssen übereinstimmende Regeln gelten. Die Internationale Kommis- sion für Zoologische Nomenklatur (ICZN) wurde im September 1895 in Leiden gegründet. -

Leiostyla Colvillei
The IUCN Red List of Threatened Species™ ISSN 2307-8235 (online) IUCN 2008: T171099A6799136 Leiostyla colvillei Assessment by: Seddon, M.B. View on www.iucnredlist.org Citation: Seddon, M.B. 2013. Leiostyla colvillei. The IUCN Red List of Threatened Species 2013: e.T171099A6799136. http://dx.doi.org/10.2305/IUCN.UK.2011-1.RLTS.T171099A6799136.en Copyright: © 2015 International Union for Conservation of Nature and Natural Resources Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale, reposting or other commercial purposes is prohibited without prior written permission from the copyright holder. For further details see Terms of Use. The IUCN Red List of Threatened Species™ is produced and managed by the IUCN Global Species Programme, the IUCN Species Survival Commission (SSC) and The IUCN Red List Partnership. The IUCN Red List Partners are: BirdLife International; Botanic Gardens Conservation International; Conservation International; Microsoft; NatureServe; Royal Botanic Gardens, Kew; Sapienza University of Rome; Texas A&M University; Wildscreen; and Zoological Society of London. If you see any errors or have any questions or suggestions on what is shown in this document, please provide us with feedback so that we can correct or extend the information provided. THE IUCN RED LIST OF THREATENED SPECIES™ Taxonomy Kingdom Phylum Class Order Family Animalia Mollusca Gastropoda Stylommatophora Lauriidae Taxon Name: Leiostyla colvillei Seddon & Killeen, 1996 Assessment Information Red List Category & Criteria: Vulnerable D2 ver 3.1 Year Published: 2013 Date Assessed: August 31, 2010 Justification: This species is endemic to Madeira, where it is only known from the type locality in the valley of Ribeira de Faja da Nogueira which lies within a remote, relatively inaccessible area. -

European Red List of Non-Marine Molluscs Annabelle Cuttelod, Mary Seddon and Eike Neubert
European Red List of Non-marine Molluscs Annabelle Cuttelod, Mary Seddon and Eike Neubert European Red List of Non-marine Molluscs Annabelle Cuttelod, Mary Seddon and Eike Neubert IUCN Global Species Programme IUCN Regional Office for Europe IUCN Species Survival Commission Published by the European Commission. This publication has been prepared by IUCN (International Union for Conservation of Nature) and the Natural History of Bern, Switzerland. The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN, the Natural History Museum of Bern or the European Union concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The views expressed in this publication do not necessarily reflect those of IUCN, the Natural History Museum of Bern or the European Commission. Citation: Cuttelod, A., Seddon, M. and Neubert, E. 2011. European Red List of Non-marine Molluscs. Luxembourg: Publications Office of the European Union. Design & Layout by: Tasamim Design - www.tasamim.net Printed by: The Colchester Print Group, United Kingdom Picture credits on cover page: The rare “Hélice catalorzu” Tacheocampylaea acropachia acropachia is endemic to the southern half of Corsica and is considered as Endangered. Its populations are very scattered and poor in individuals. This picture was taken in the Forêt de Muracciole in Central Corsica, an occurrence which was known since the end of the 19th century, but was completely destroyed by a heavy man-made forest fire in 2000. -

Predicting the Impacts of Climate Change on the Distribution and Conservation of Endemic Forest Land Snails of Madeira Island
Predicting the impacts of climate change on the distribution and conservation of endemic forest land snails of Madeira Island Cátia Sofia Alves Gouveia Predicting the impacts of climate change on the distribution and conservation of endemic forest land snails of Madeira Island Dissertation supervised by Pedro Cardoso, PhD Finnish Museum of Natural History, University of Helsinki Robert A. D. Cameron, PhD The University of Sheffield November 2013 ii ACKNOWLEDGEMENTS First of all, I would like to thank my supervisors Pedro Cardoso and Robert A. D. Cameron, for the supervision and support given in the development of this work. I am extremely grateful for the opportunity to be under their guidance, for all their inspiration and scientific support during the different stages of this work. I am convinced that my future as a researcher will be greatly influenced by their experience and knowledge. I would also like to thank Dinarte Teixeira, for all the support given since the draft of the first ideas of this project and for sharing his knowledge and enthusiasm about modeling, land snails and conservation. His commitment to this work and valuable contributions were crucial during the entire project. I am also extremely grateful to Direção Regional do Ordenamento do Território e Ambiente, for their support and access to the BIOBASE, and also for providing digital cartography and geographic information. A special thank to the Centre for Climate Change Impacts Adaptation & Modeling, who kindly provided future climate scenarios, modelled under the CLIMAAT II project (Climate and Meteorology of the Atlantic Islands (Azores, Madeira and Canary Islands)). I owe a special debt of appreciation to the Natural History Museum of London, for allowing the examination of molluscan collection from Madeira Island. -

Zoologische Mededelingen
ZOOLOGISCHE MEDEDELINGEN UITGEGEVEN DOOR HET RIJKSMUSEUM VAN NATUURLIJKE HISTORIE TE LEIDEN (MINISTERIE VAN WELZIJN, VOLKSGEZONDHEID EN CULTUUR) Deel 57 no. 23 15 december 1983 ON IBERIAN COCHLICOPIDAE AND THE GENUS CRYPTA ZECA (GASTROPODA, PULMONATA) by E. GITTENBERGER Rijksmuseum van Natuurlijke Historie, Leiden, The Netherlands With 13 text-figures ABSTRACT Some new data concerning the four species of Cochlicopidae known from the Iberian peninsu- la, viz., Azeca goodalli, Hypnophila boissii, Cochlicopa lubrica, and C. lubricella, are presented. In addition, Hypnophila malagana spec. nov. is described (by Gittenberger & Menkhorst). Cochlicopa lubrica (Müller, 1774) is designated (by Gittenberger & Waldén) as the type-species of Cochlicopa Férussac, April 1821. It is shown that the genus name Cionella Jeffreys, 1829, should not be used instead of Cochlicopa Férussac, April 1821. The genus Cryptazeca, still of uncertain systematic position, but conchologically similar to the Cochlicopidae, is revised. Anatomical characters could not be used. The data concerning Crypta- zeca monodonta, C. subcylindrica, and C. vasconica are summarized. In addition C. kobelti spec. nov. is described. INTRODUCTION Four species of Cochlicopidae are known to occur on the Iberian peninsu• la; a fifth species is described as new in the present paper. Distribution maps (50 km UTM-grid) are given for all species; these may cover the entire range of the species or only the Iberian part of it. The locality data in the text are provided with the 10 km or 1 km UTM-grid code. In addition to the distribu• tional data, various notes are given, depending on the state of our knowledge concerning the species in question.