NOTES on the DISARTICULATION of XYLOTHAMIA (ASTERACEAE: ASTEREAE) Guy L
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Revised Section 4.6 Biological Resources Draft Environmental
Revised Section 4.6 Biological Resources Draft Environmental Impact Report SCH No. 2002091081 Prepared for: City of Santa Clarita Department of Planning & Building Services 23920 Valencia Boulevard, Suite 302 Santa Clarita, California 91355 Prepared by: Impact Sciences, Inc. 30343 Canwood Street, Suite 210 Agoura Hills, California 91301 March 2004 Revised Section 4.6 Biological Resources Draft Environmental Impact Report SCH No. 2002091081 Prepared for: City of Santa Clarita Department of Planning & Building Services 23920 Valencia Boulevard, Suite 302 Santa Clarita, California 91355 Prepared by: Impact Sciences, Inc. 30343 Canwood Street, Suite 210 Agoura Hills, California 91301 March 2004 Table of Contents Volume I: Environmental Impact Report Section Page Introduction....................................................................................................................I-1 Executive Summary ......................................................................................................ES-1 1.0 Project Description....................................................................................................... 1.0-1 2.0 Environmental and Regulatory Setting......................................................................... 2.0-1 3.0 Cumulative Impact Analysis Methodology .................................................................. 3.0-1 4.0 Environmental Impact Analyses................................................................................... 4.0-1 4.1 Geotechnical Hazards.......................................................................................... -

Tejon Ranch Botanical Survey Report
David Magney Environmental Consulting FLORA OF THE TEJON RANCH CONSERVANCY ACQUISITION AREAS, TEJON RANCH, CALIFORNIA Prepared for: TEJON RANCH CONSERVANCY July 2010 DMEC Mission Statement: To provide quality environmental consulting services, with integrity, that protect and enhance the human and natural environment. David Magney Environmental Consulting Flora of the Tejon Ranch Conservancy Acquisition Areas, Tejon Ranch, California Prepared for: Tejon Ranch Conservancy P.O. Box 216 Frazier Park, California 93225 Contact: Michael White Phone: 661/-248-2400 ext 2 Prepared by: David Magney Environmental Consulting P.O. Box 1346 Ojai, California 93024-1346 Phone: 805/646-6045 23 July 2010 DMEC Mission Statement: To provide quality environmental consulting services, with integrity, that protect and enhance the human and natural environment. This document should be cited as: David Magney Environmental Consulting. 2010. Flora of the Tejon Ranch Conservancy Acquisition Areas, Tejon Ranch, California. 23 July2010. (PN 09-0001.) Ojai, California. Prepared for Tejon Ranch Conservancy, Frazier Park, California. Tejon Ranch Conservancy – Flora of Tejon Ranch Acquisition Areas Project No. 09-0001 DMEC July 2010 TABLE OF CONTENTS Page SECTION 1. INTRODUCTION............................................................................. 1 SECTION 2. METHODS ........................................................................................ 3 Field Survey Methods .......................................................................................................... -

2014 Plant Species List
Acanthaceae Hygrophila Occasional lacustris Acanthaceae Justicia ovata Uncommon Acanthaceae Ruellia humilis Common Acanthaceae Ruellia nudiflora s.n. Uncommon Acanthaceae Ruellia Occasional pedunculata Aceraceae Acer rubrum Occasional Agavaceae Yucca louisianica Uncommon Aiozaceae Molluga Occasional verticillata Alismataceae Echinodorus Occasional cordifolius Alismataceae Sagittaria Rare papillosa Alismataceae Sagittaria 156 Uncommon platyphylla Alliaceae Allium Occasional canadense var. canadense Alliaceae Allium Occasional canadense var. mobilense Alliaceae Allium 96, Uncommon drummondii 124 (Keith 96, 124) Amaranthaceae Alternanthera Common philoxeroides Amaryllidaceae Hymenocallis Uncommon liriosome Anacardiaceae Rhus aromatica Uncommon Anacardiaceae Rhus copallinum Occasional Anacardiaceae Toxicodendron Frequent radicans Apiaceae Bifora americana Common Apiaceae Centella erecta Uncommon Apiaceae Chaerophyllum Uncommon tainturieri Apiaceae Cicuta Uncommon maculatum Apiaceae Cynosciadium Uncommon digitatum Apiaceae Eryngium Common yuccifolium Apiaceae Hydrocotyle Occasional verticillata Apiaceae Polytaenia Frequent texana Apiaceae Ptilimnium Common capillaceum Apiaceae Ptilimnium Common nuttallii Apiaceae Spermolepsis Common inermis Apiaceae Torilis arvensis Occasional Apocynaceae Apocynum Occasional cannibinum Apocynaceae Nerium oleander Rare Apocynaceae Trachelospermu Occasional m difforme Aquifoliaceae Ilex decidua Common Aquifoliaceae Ilex opaca Common Aquifoliaceae Ilex vomitoria Abundant Araceae Arisaema Rare dracontium Araceae -

Biological Resources Report for the Bella Terrazza Project, El Cajon, California
Biological Resources Report for the Bella Terrazza Project, El Cajon, California Prepared for Prepared by Priest Development Corporation RECON Environmental, Inc. 124 West Main Street, Suite 240 1927 Fifth Avenue El Cajon, CA 92020 San Diego, CA 92101-2358 Contact: Asher Burke P 619.308.9333 F 619.308.9334 RECON Number 7774 March 26, 2015 Kevin Israel, Biologist THIS PAGE IS INTENTIONALLY BLANK. Biological Resources Report for the Bella Terrazza Project TABLE OF CONTENTS 1.0 Summary 1 2.0 Introduction 1 3.0 Survey Methods 5 4.0 Existing Conditions 5 4.1 Botanical Resources 5 4.2 Zoological Resources 10 4.3 Sensitive Biological Resources 10 5.0 Project Impacts 12 5.1 Sensitive Plants 12 5.2 Sensitive Wildlife 12 6.0 Mitigation 14 7.0 References Cited 14 FIGURES 1: Regional Location 2 2: Project Location on USGS Map 3 3: Project Location on Aerial Photograph 4 4: Existing Land Cover Types 6 5: Impacts to Biological Resources 13 TABLES 1: Existing Land Cover Types within the Survey Area 10 2: Impacts to Land Cover Types 12 PHOTOGRAPHS 1: View of Urban/Developed Land and Disturbed Land Looking Northeast from East Main Street 7 2: View of Disturbed Land Looking East from East Main Street 7 3: View of Ditch Looking North along East Main Street 8 4: View of Disturbed Land Looking East along the Northern Boundary 8 5: View of Disturbed Land Looking South along the Eastern Boundary 9 ATTACHMENTS 1: Plant Species Observed Within the Bella Terrazza Project Site 2: Wildlife Species Observed/Detected Within the Bella Terrazza Project Site 3: Sensitive Plant Species Observed or with the Potential to Occur on the Bella Terrazza Project Site 4: Sensitive Wildlife Species Observed or with the Potential to Occur on the Bella Terrazza Project Site Biological Resources Report for the Bella Terrazza Project THIS PAGE IS INTENTIONALLY BLANK. -

Literature Cited
Literature Cited Robert W. Kiger, Editor This is a consolidated list of all works cited in volumes 19, 20, and 21, whether as selected references, in text, or in nomenclatural contexts. In citations of articles, both here and in the taxonomic treatments, and also in nomenclatural citations, the titles of serials are rendered in the forms recommended in G. D. R. Bridson and E. R. Smith (1991). When those forms are abbre- viated, as most are, cross references to the corresponding full serial titles are interpolated here alphabetically by abbreviated form. In nomenclatural citations (only), book titles are rendered in the abbreviated forms recommended in F. A. Stafleu and R. S. Cowan (1976–1988) and F. A. Stafleu and E. A. Mennega (1992+). Here, those abbreviated forms are indicated parenthetically following the full citations of the corresponding works, and cross references to the full citations are interpolated in the list alphabetically by abbreviated form. Two or more works published in the same year by the same author or group of coauthors will be distinguished uniquely and consistently throughout all volumes of Flora of North America by lower-case letters (b, c, d, ...) suffixed to the date for the second and subsequent works in the set. The suffixes are assigned in order of editorial encounter and do not reflect chronological sequence of publication. The first work by any particular author or group from any given year carries the implicit date suffix “a”; thus, the sequence of explicit suffixes begins with “b”. Works missing from any suffixed sequence here are ones cited elsewhere in the Flora that are not pertinent in these volumes. -

Texas Prairie Dawn-Flower (Hymenoxys Texana) 5-Year Review
Texas prairie dawn-flower (Hymenoxys texana) 5-Year Review: Summary and Evaluation Photo credit: USFWS U.S. Fish and Wildlife Service Texas Coastal Ecological Services Field Office Houston, Texas Table of Contents ABBREVIATIONS ........................................................................................................................ 3 1.0 GENERAL INFORMATION .......................................................................................... 4 2.0 REVIEW ANALYSIS ...................................................................................................... 7 2.4 SYNTHESIS .................................................................................................................. 24 3.0 RESULTS....................................................................................................................... 25 4.0 RECOMMENDATIONS FOR FUTURE ACTIONS.................................................... 26 5.0 REFERENCES ............................................................................................................... 28 Appendix A ................................................................................................................................... 31 Recommendation resulting from the 5-Year Review: .................................................................. 34 Figures Figure 1 Current H. texana county occurrences ............................................................................. 9 Tables Table 1 Renaming of species historically associated with H. texana .......................................... -
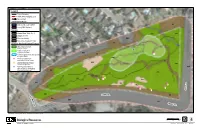
Biological Resources 0 50 100 I FIGURE 4 SKYLINE RETIREMENT CENTER Aerial Source: Google Earth, Nov
Legend Project Boundary 100-ft Off-site Mapping Limit Survey Route Jurisdictional Areas Waters of the U.S. / CDFW DIS County RPO Wetland Habitats CSS Coastal Sage Scrub (32510) CSS CSS DIS Disturbed (11300) DEV Developed (12000) DEV NNG Non-native Grassland (42200) Sensitive Species & Unique Features DIS San Diego Sunflower Bahiopsis laciniata ! Coast Cholla patch Cylindropuntia prolifera DEV DIS California gnatcatcher activity area 12 Polioptila californica CSS ! Palmer's Goldenbush Ericameria palmeri var. palm eri ! NNG ! Orange-throated Whiptail CSS Aspidoscelis hyperythra ! Disintegrated midden of DEV San Diego Desert Woodrat Neotoma lepida intermedia DIS CSS DIS 1 CSS 10 3 NNG DEV NNG 11 NNG 1 DIS 2 1 1 RD PRIVATE CSS CAMPO RD. DIS V IA M DIS E DEV R C DEV A D C AMPO O RD. T:\Project_Data\Skyline_Retirement_Center\Final_Maps\CAGN_Report_050817\SC_Fig-04_BioResMap_051017.mxd Feet Biological Resources 0 50 100 I FIGURE 4 SKYLINE RETIREMENT CENTER Aerial Source: Google Earth, Nov. 2016 May 2017 APPENDIX G 2017 California Gnatcatcher Survey Report for the Skyline Retirement Western Offsite Mitigation Parcel REC Consultants, Inc. Skyline Retirement Center July 2018 Biological Resources Letter Report May 19, 2017 Stacey Love Recovery Permit Coordinator U.S. Fish and Wildlife Service Carlsbad Fish and Wildlife Office 2177 Salk Avenue, Suite 250 Carlsbad, CA 92008 Subject: 2017 California Gnatcatcher Survey Report for the Skyline Retirement Center Western Offsite Mitigation Parcel, San Diego County, California; APN 506-140-08; USFWS Permit TE786714-1 Ms. Love: This report provides the results of a protocol survey series for the coastal California gnatcatcher (Polioptila californica californica) performed by REC biologists Elyssa Robertson and Catherine MacGregor on the western offsite mitigation parcel associated with the Skyline Retirement Center project. -

References and Appendices
References Ainley, D.G., S.G. Allen, and L.B. Spear. 1995. Off- Arnold, R.A. 1983. Ecological studies on six endan- shore occurrence patterns of marbled murrelets gered butterflies (Lepidoptera: Lycaenidae): in central California. In: C.J. Ralph, G.L. Hunt island biogeography, patch dynamics, and the Jr., M.G. Raphael, and J.F. Piatt, technical edi- design of habitat preserves. University of Cali- tors. Ecology and Conservation of the Marbled fornia Publications in Entomology 99: 1–161. Murrelet. USDA Forest Service, General Techni- Atwood, J.L. 1993. California gnatcatchers and coastal cal Report PSW-152; 361–369. sage scrub: the biological basis for endangered Allen, C.R., R.S. Lutz, S. Demairais. 1995. Red im- species listing. In: J.E. Keeley, editor. Interface ported fire ant impacts on Northern Bobwhite between ecology and land development in Cali- populations. Ecological Applications 5: 632-638. fornia. Southern California Academy of Sciences, Allen, E.B., P.E. Padgett, A. Bytnerowicz, and R.A. Los Angeles; 149–169. Minnich. 1999. Nitrogen deposition effects on Atwood, J.L., P. Bloom, D. Murphy, R. Fisher, T. Scott, coastal sage vegetation of southern California. In T. Smith, R. Wills, P. Zedler. 1996. Principles of A. Bytnerowicz, M.J. Arbaugh, and S. Schilling, reserve design and species conservation for the tech. coords. Proceedings of the international sym- southern Orange County NCCP (Draft of Oc- posium on air pollution and climate change effects tober 21, 1996). Unpublished manuscript. on forest ecosystems, February 5–9, 1996, River- Austin, M. 1903. The Land of Little Rain. University side, CA. -

Reclassification of North American Haplopappus (Compositae: Astereae) Completed: Rayjacksonia Gen
AmericanJournal of Botany 83(3): 356-370. 1996. RECLASSIFICATION OF NORTH AMERICAN HAPLOPAPPUS (COMPOSITAE: ASTEREAE) COMPLETED: RAYJACKSONIA GEN. NOV.1 MEREDITH A. LANE2 AND RONALD L. HARTMAN R. L. McGregor Herbarium(University of Kansas NaturalHistory Museum Division of Botany) and Departmentof Botany,University of Kansas, Lawrence, Kansas 66047-3729; and Rocky MountainHerbarium, Department of Botany,University of Wyoming,Laramie, Wyoming82071-3165 Rayjacksonia R. L. Hartman& M. A. Lane, gen. nov. (Compositae: Astereae), is named to accommodate the "phyllo- cephalus complex," formerlyof Haplopappus Cass. sect. Blepharodon DC. The new combinationsare R. phyllocephalus (DC.) R. L. Hartman& M. A. Lane, R. annua (Rydb.) R. L. Hartman& M. A. Lane, and R. aurea (A. Gray) R. L. Hartman & M. A. Lane. This transfercompletes the reclassificationof the North American species of Haplopappus sensu Hall, leaving that genus exclusively South American.Rayjacksonia has a base chromosomenumber of x = 6. Furthermore,it shares abruptlyampliate disk corollas, deltatedisk style-branchappendages, and corolla epidermalcell type,among other features,with Grindelia, Isocoma, Olivaea, Prionopsis, Stephanodoria, and Xanthocephalum.Phylogenetic analyses of morphologicaland chloroplastDNA restrictionsite data, taken together,demonstrate that these genera are closely related but distinct. Key words: Astereae; Asteraceae; Compositae; Haplopappus; Rayjacksonia. During the past seven decades, taxonomic application lopappus sensu Hall (1928) are reclassifiedand are cur- -

Groundcover Restoration in Forests of the Southeastern United States
Groundcover RestorationIN FORESTS OF THE SOUTHEASTERN UNITED STATES Jennifer L. Trusty & Holly K. Ober Acknowledgments The funding for this project was provided by a cooperative • Florida Fish and Wildlife Conservation Commission of resource managers and scientific researchers in Florida, • Florida Department of Environmental Protection Conserved Forest Ecosystems: Outreach and Research • Northwest Florida Water Management District (CFEOR). • Southwest Florida Water Management District • Suwannee River Water Management District CFEOR is a cooperative comprised of public, private, non- government organizations, and landowners that own or We are grateful to G. Tanner for making the project manage Florida forest lands as well as University of Florida possible and for providing valuable advice on improving the faculty members. CFEOR is dedicated to facilitating document. We are also indebted to the many restorationists integrative research and outreach that provides social, from across the Southeast who shared information with J. ecological, and economic benefits to Florida forests on a Trusty. Finally, we thank H. Kesler for assistance with the sustainable basis. Specifically, funding was provided by maps and L. DeGroote, L. Demetropoulos, C. Mackowiak, C. Matson and D. Printiss for assistance with obtaining photographs. Cover photo: Former slash pine plantation with restored native groundcover. Credits: L. DeGroote. Suggested citation: Trusty, J. L., and H. K. Ober. 2009. Groundcover restoration in forests of the Southeastern United States. CFEOR Research Report 2009-01. University of Florida, Gainesville, FL. 115 pp. | 3 | Table of Contents INTRODUCTION . 7 PART I - Designing and Executing a Groundcover PART II – Resources to Help Get the Job Done Restoration Project CHAPTER 6: Location of Groundcover CHAPTER 1: Planning a Restoration Project . -

Flora of the Carolinas, Virginia, and Georgia, Working Draft of 17 March 2004 -- BIBLIOGRAPHY
Flora of the Carolinas, Virginia, and Georgia, Working Draft of 17 March 2004 -- BIBLIOGRAPHY BIBLIOGRAPHY Ackerfield, J., and J. Wen. 2002. A morphometric analysis of Hedera L. (the ivy genus, Araliaceae) and its taxonomic implications. Adansonia 24: 197-212. Adams, P. 1961. Observations on the Sagittaria subulata complex. Rhodora 63: 247-265. Adams, R.M. II, and W.J. Dress. 1982. Nodding Lilium species of eastern North America (Liliaceae). Baileya 21: 165-188. Adams, R.P. 1986. Geographic variation in Juniperus silicicola and J. virginiana of the Southeastern United States: multivariant analyses of morphology and terpenoids. Taxon 35: 31-75. ------. 1995. Revisionary study of Caribbean species of Juniperus (Cupressaceae). Phytologia 78: 134-150. ------, and T. Demeke. 1993. Systematic relationships in Juniperus based on random amplified polymorphic DNAs (RAPDs). Taxon 42: 553-571. Adams, W.P. 1957. A revision of the genus Ascyrum (Hypericaceae). Rhodora 59: 73-95. ------. 1962. Studies in the Guttiferae. I. A synopsis of Hypericum section Myriandra. Contr. Gray Herbarium Harv. 182: 1-51. ------, and N.K.B. Robson. 1961. A re-evaluation of the generic status of Ascyrum and Crookea (Guttiferae). Rhodora 63: 10-16. Adams, W.P. 1973. Clusiaceae of the southeastern United States. J. Elisha Mitchell Sci. Soc. 89: 62-71. Adler, L. 1999. Polygonum perfoliatum (mile-a-minute weed). Chinquapin 7: 4. Aedo, C., J.J. Aldasoro, and C. Navarro. 1998. Taxonomic revision of Geranium sections Batrachioidea and Divaricata (Geraniaceae). Ann. Missouri Bot. Gard. 85: 594-630. Affolter, J.M. 1985. A monograph of the genus Lilaeopsis (Umbelliferae). Systematic Bot. Monographs 6. Ahles, H.E., and A.E. -

Illustrated Flora of East Texas Illustrated Flora of East Texas
ILLUSTRATED FLORA OF EAST TEXAS ILLUSTRATED FLORA OF EAST TEXAS IS PUBLISHED WITH THE SUPPORT OF: MAJOR BENEFACTORS: DAVID GIBSON AND WILL CRENSHAW DISCOVERY FUND U.S. FISH AND WILDLIFE FOUNDATION (NATIONAL PARK SERVICE, USDA FOREST SERVICE) TEXAS PARKS AND WILDLIFE DEPARTMENT SCOTT AND STUART GENTLING BENEFACTORS: NEW DOROTHEA L. LEONHARDT FOUNDATION (ANDREA C. HARKINS) TEMPLE-INLAND FOUNDATION SUMMERLEE FOUNDATION AMON G. CARTER FOUNDATION ROBERT J. O’KENNON PEG & BEN KEITH DORA & GORDON SYLVESTER DAVID & SUE NIVENS NATIVE PLANT SOCIETY OF TEXAS DAVID & MARGARET BAMBERGER GORDON MAY & KAREN WILLIAMSON JACOB & TERESE HERSHEY FOUNDATION INSTITUTIONAL SUPPORT: AUSTIN COLLEGE BOTANICAL RESEARCH INSTITUTE OF TEXAS SID RICHARDSON CAREER DEVELOPMENT FUND OF AUSTIN COLLEGE II OTHER CONTRIBUTORS: ALLDREDGE, LINDA & JACK HOLLEMAN, W.B. PETRUS, ELAINE J. BATTERBAE, SUSAN ROBERTS HOLT, JEAN & DUNCAN PRITCHETT, MARY H. BECK, NELL HUBER, MARY MAUD PRICE, DIANE BECKELMAN, SARA HUDSON, JIM & YONIE PRUESS, WARREN W. BENDER, LYNNE HULTMARK, GORDON & SARAH ROACH, ELIZABETH M. & ALLEN BIBB, NATHAN & BETTIE HUSTON, MELIA ROEBUCK, RICK & VICKI BOSWORTH, TONY JACOBS, BONNIE & LOUIS ROGNLIE, GLORIA & ERIC BOTTONE, LAURA BURKS JAMES, ROI & DEANNA ROUSH, LUCY BROWN, LARRY E. JEFFORDS, RUSSELL M. ROWE, BRIAN BRUSER, III, MR. & MRS. HENRY JOHN, SUE & PHIL ROZELL, JIMMY BURT, HELEN W. JONES, MARY LOU SANDLIN, MIKE CAMPBELL, KATHERINE & CHARLES KAHLE, GAIL SANDLIN, MR. & MRS. WILLIAM CARR, WILLIAM R. KARGES, JOANN SATTERWHITE, BEN CLARY, KAREN KEITH, ELIZABETH & ERIC SCHOENFELD, CARL COCHRAN, JOYCE LANEY, ELEANOR W. SCHULTZE, BETTY DAHLBERG, WALTER G. LAUGHLIN, DR. JAMES E. SCHULZE, PETER & HELEN DALLAS CHAPTER-NPSOT LECHE, BEVERLY SENNHAUSER, KELLY S. DAMEWOOD, LOGAN & ELEANOR LEWIS, PATRICIA SERLING, STEVEN DAMUTH, STEVEN LIGGIO, JOE SHANNON, LEILA HOUSEMAN DAVIS, ELLEN D.