Reinterpretation of Hinge Structures of Two Devonian Bivalvia: Congeriomorpha and Tvsayan A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Field Identification Guide to the Living Marine Resources In
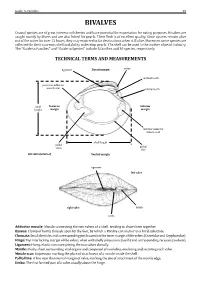
Guide to Families 29 BIVALVES Coastal species are of great interest to fisheries and have potential for exportation for eating purposes. Bivalves are caught mainly by divers and are also fished for pearls. Their flesh is of excellent quality. Since oysters remain alive out of the water for over 12 hours, they may exported to far destinations when still alive. Moreover, some species are collected for their nacreous shell and ability to develop pearls. The shell can be used in the mother of pearl industry. The “Guide to Families’’ andTECHNICAL ‘‘Guide to Species’’ TERMS include 5AND families MEASUREMENTS and 10 species, respectively. ligament Dorsal margin umbo posterior adductor cardinal tooth muscle scar lateral tooth Posterior Anterior margin margin shell height anterior adductor muscle scar pallial sinus pallial shell length line left valve (interior) Ventral margin ligament left valve right valve lunule umbo Adductor muscle: Byssus: Chomata: Muscle connecting the two valves of a shell, tending to draw them together. Hinge: Clump of horny threads spun by the foot, by which a Bivalve can anchor to a hard substrate. Ligament: Small denticles and corresponding pits located on the inner margin of the valves (Ostreidae and Gryphaeidae). Mantle: Top interlocking margin of the valves, often with shelly projections (teeth) and corresponding recesses (sockets). Muscle scar: Horny, elastic structure joining the two valves dorsally. Pallial line: Fleshy sheet surrounding vital organs and composed of two lobes, one lining and secreting each valve. Umbo: Impression marking the place of attachment of a muscle inside the shell. A line near the internal margin of valve, marking the site of attachment of the mantle edge. -

Freshwater Mussels of the Pacific Northwest
Freshwater Mussels of the Pacifi c Northwest Ethan Nedeau, Allan K. Smith, and Jen Stone Freshwater Mussels of the Pacifi c Northwest CONTENTS Part One: Introduction to Mussels..................1 What Are Freshwater Mussels?...................2 Life History..............................................3 Habitat..................................................5 Role in Ecosystems....................................6 Diversity and Distribution............................9 Conservation and Management................11 Searching for Mussels.............................13 Part Two: Field Guide................................15 Key Terms.............................................16 Identifi cation Key....................................17 Floaters: Genus Anodonta.......................19 California Floater...................................24 Winged Floater.....................................26 Oregon Floater......................................28 Western Floater.....................................30 Yukon Floater........................................32 Western Pearlshell.................................34 Western Ridged Mussel..........................38 Introduced Bivalves................................41 Selected Readings.................................43 www.watertenders.org AUTHORS Ethan Nedeau, biodrawversity, www.biodrawversity.com Allan K. Smith, Pacifi c Northwest Native Freshwater Mussel Workgroup Jen Stone, U.S. Fish and Wildlife Service, Columbia River Fisheries Program Offi ce, Vancouver, WA ACKNOWLEDGEMENTS Illustrations, -

FAO Fisheries & Aquaculture
Food and Agriculture Organization of the United Nations Fisheries and for a world without hunger Aquaculture Department Cultured Aquatic Species Information Programme Mercenaria mercenaria (Linnaeus, 1758) I. Identity V. Status And Trends a. Biological Features VI. Main Issues b. Images Gallery a. Responsible Aquaculture Practices II. Profile VII. References a. Historical Background a. Related Links b. Main Producer Countries c. Habitat And Biology III. Production a. Production Cycle b. Production Systems c. Diseases And Control Measures IV. Statistics a. Production Statistics b. Market And Trade Identity Mercenaria mercenaria Linnaeus, 1758 [Veneridae] FAO Names: En - Northern quahog(=Hard clam), Fr - Praire, Es - Chirla mercenaria Biological features Shell solid, equivalve; inequilateral, beaks in the front half of the shell; broadly oval in outline. Ligament a deeply inset, dark brown elliptical band, behind the beaks reaching half-way to the posterior margin. Lunule well defined, broad, heart-shaped. Escutcheon indistinct. Sculpture of concentric lines, raised here and there into ridges, and fine radiating lines. In young specimens the ridges are present all over the shell but in the adult they persist, after wear and tear, only near the anterior and posterior margins. Growth stages prominent. Both valves with three cardinal teeth; in addition there is present in each valve a rough tooth-like area behind the beaks and immediately below the ligament; this area has the appearance of a supplementary posterior cardinal tooth which has been broken off. No laterals. Pallial sinus not deep, triangular. Margin grenulate. Colour a dirty white, light varnish-brown, dull grey or grey-brown. Inside of shell white, sometimes deep violet about the adductor muscle scars. -

Key to the Freshwater Bivalves of New Jersey
Key to the Freshwater Bivalves of New Jersey 1. a. shell with a very sharp posterior ridge, shaped like the marine mussel, Mytilus, generally less than 30 mm, and attached to a hard substrate with byssal threads.........................……………........................Zebra mussel b. animal without byssal threads attaching adult to substrate, with or without teeth but not with the above shape................................….............................2 2. a. valves with cardinal teeth and two sets of lateral teeth.......................…...............................3 b. valves with one set of lateral teeth and pseudocardinal teeth or without teeth.............................................................................................................5 3. a. shell thick and sturdy, beak bulbous and curving anteriorly………………….Atlantic rangia b. shell moderately thick, beak not bulbous nor curving…………………………………………...4 4. a. valves with serrated lateral teeth......................................……….........................Asian clam b. valves with smooth lateral teeth....................................................................Fingernail clam 5. a. hinge teeth absent.................................................................................................................6 b. hinge teeth present..............................................................................................................10 6. a. beaks not projecting above the hinge line................…………………........ Paper pondshell b. beaks projecting above -

Transposed Hinge Structures in Lamellibranches
TRANSACTIONS OF THE • SAN DIEGO SOCIETY OF NATURAL HISTORY ,/ VoLUME VII, No. 26, pp. 299-318, plate 19 • TRANSPOSED HINGE STRUCTURES IN LAMELLIBRANCHS BY P. PoPENOE AND FINDLAY W. W. A. • Balch Graduate School of the Geological Sciences California Institute of Technology Pasadena, California SAN DIEGO, CALIFORNIA PRINTED FOR THE SOCIETY OCTOBER 6, 1933 • • COMMITTEE ON PUBLICATION U.S. GRANT, IV, Chairman FRED BAKER CLINTON G. ABBOTT, Editor • TRANSPOSED HINGE STRUCTURES IN LAMELLIBRANCHS 1 BY W. P. PoPENOE AND W. A. FINDLAY California Institute of Technology INTRODUCTION In the course of study of a collection of Eocene fossils from Claiborne Bluff, Alabama, the senior author of tl1is paper noticed two valves, one right and one left, of the lamellibranch Venericardia parva Lea, in whicl1 the dentition is partially transposed. Subsequently, the authors made an examination of more than five thousand lamellibranch valves, representit1g both recent and fossil shells, in search of further exa1nples of hinge-trans position. We have found a total number of twenty-six valves exhibiting this variation. Study of these specimens has revealed some hitherto unreported facts regarding the principles of hinge-transposition. Therefore in this paper, we shall describe and discuss these specimens, and shall present such con clusions as seem justified by the data assembled. Citations to the literature are made by author, date, and page, referring to the list at the end of the paper. DEFINITION OF T ERMS A transposed lamellibranch hinge is defined as one that exhibits in the right valve the hinge ele1nents normally occurring in the left valve, and vice-versa. -

Mechanics Unlocks the Morphogenetic Puzzle of Interlocking Bivalved Shells
Mechanics unlocks the morphogenetic puzzle of interlocking bivalved shells Derek E. Moultona,1 , Alain Gorielya , and Regis´ Chiratb aMathematical Institute, University of Oxford, Oxford, OX2 6GG, United Kingdom; and bCNRS 5276, LGL-TPE (Le Laboratoire de Geologie´ de Lyon: Terre, Planetes,` Environnement), Universite´ Lyon 1, 69622 Villeurbanne Cedex, France Edited by Sean H. Rice, Texas Tech University, Lubbock, TX, and accepted by Editorial Board Member David Jablonski November 11, 2019 (received for review September 24, 2019) Brachiopods and mollusks are 2 shell-bearing phyla that diverged tal events causing shell injuries. Yet, in all cases the interlocking from a common shell-less ancestor more than 540 million years ago. of the 2 shell edges is tightly maintained. These observations Brachiopods and bivalve mollusks have also convergently evolved imply that the interlocking pattern emerges as the result of epi- a bivalved shell that displays an apparently mundane, yet strik- genetic interactions modulating the behavior of the secreting ing feature from a developmental point of view: When the shell mantle during shell development. is closed, the 2 valve edges meet each other in a commissure that Here, we provide a geometric and mechanical explanation forms a continuum with no gaps or overlaps despite the fact that for this morphological trait based on a detailed analysis of the each valve, secreted by 2 mantle lobes, may present antisymmet- shell geometry during growth and the physical interaction of the ric ornamental patterns of varying regularity and size. Interlock- shell-secreting soft mantle with both the rigid shell edge and ing is maintained throughout the entirety of development, even the opposing mantle lobe. -

Conceptual Issues in Phylogeny, Taxonomy, and Nomenclature
Contributions to Zoology, 66 (1) 3-41 (1996) SPB Academic Publishing bv, Amsterdam Conceptual issues in phylogeny, taxonomy, and nomenclature Alexandr P. Rasnitsyn Paleontological Institute, Russian Academy ofSciences, Profsoyuznaya Street 123, J17647 Moscow, Russia Keywords: Phylogeny, taxonomy, phenetics, cladistics, phylistics, principles of nomenclature, type concept, paleoentomology, Xyelidae (Vespida) Abstract On compare les trois approches taxonomiques principales développées jusqu’à présent, à savoir la phénétique, la cladis- tique et la phylistique (= systématique évolutionnaire). Ce Phylogenetic hypotheses are designed and tested (usually in dernier terme s’applique à une approche qui essaie de manière implicit form) on the basis ofa set ofpresumptions, that is, of à les traits fondamentaux de la taxonomic statements explicite représenter describing a certain order of things in nature. These traditionnelle en de leur et particulier son usage preuves ayant statements are to be accepted as such, no matter whatever source en même temps dans la similitude et dans les relations de evidence for them exists, but only in the absence ofreasonably parenté des taxons en question. L’approche phylistique pré- sound evidence pleading against them. A set ofthe most current sente certains avantages dans la recherche de réponses aux phylogenetic presumptions is discussed, and a factual example problèmes fondamentaux de la taxonomie. ofa practical realization of the approach is presented. L’auteur considère la nomenclature A is made of the three -

Exputens) in Mexico, and a Review of All Species of This North American Subgenus
Natural History Museum /U, JH caY-^A 19*90 la Of Los Angeles County THE VELIGER © CMS, Inc., 1990 The Veliger 33(3):305-316 (July 2, 1990) First Occurrence of the Tethyan Bivalve Nayadina (.Exputens) in Mexico, and a Review of All Species of This North American Subgenus by RICHARD L. SQUIRES Department of Geological Sciences, California State University, Northridge, California 91330, USA Abstract. The malleid bivalve Nayadina (Exputens) has Old World Tethyan affinities but is known only from Eocene deposits in North America. Nayadina (Exputens) is reported for the first time from Mexico. About 50 specimens of N. (E.) batequensis sp. nov. were found in warm-water nearshore deposits of the middle lower Eocene part of the Bateque Formation, just south of Laguna San Ignacio, on the Pacific coast of Baja California Sur. The new species shows a wide range of morphologic variability especially where the beaks and auricles are located and how much they are developed. A review of the other species of Exputens, namely Nayadina (E.) llajasensis (Clark, 1934) from California and N. (E.) ocalensis (MacNeil, 1934) from Florida, Georgia, and North Carolina, revealed that they also have a wide range of morphologic variability. Nayadina (E.) alexi (Clark, 1934) is shown, herein, to be a junior synonym of N. (E.) llajasensis. The presence of a byssal sinus is recognized for the first time in Exputens. An epifaunal nestling mode of life, with attachment by byssus to hard substrate, can now be assumed for Exputens. INTRODUCTION species. It became necessary to thoroughly examine them, The macropaleontology of Eocene marine deposits in Baja and after such a study, it was found that the Bateque California Sur, Mexico, is largely an untouched subject. -

Atlas of the Freshwater Mussels (Unionidae)
1 Atlas of the Freshwater Mussels (Unionidae) (Class Bivalvia: Order Unionoida) Recorded at the Old Woman Creek National Estuarine Research Reserve & State Nature Preserve, Ohio and surrounding watersheds by Robert A. Krebs Department of Biological, Geological and Environmental Sciences Cleveland State University Cleveland, Ohio, USA 44115 September 2015 (Revised from 2009) 2 Atlas of the Freshwater Mussels (Unionidae) (Class Bivalvia: Order Unionoida) Recorded at the Old Woman Creek National Estuarine Research Reserve & State Nature Preserve, Ohio, and surrounding watersheds Acknowledgements I thank Dr. David Klarer for providing the stimulus for this project and Kristin Arend for a thorough review of the present revision. The Old Woman Creek National Estuarine Research Reserve provided housing and some equipment for local surveys while research support was provided by a Research Experiences for Undergraduates award from NSF (DBI 0243878) to B. Michael Walton, by an NOAA fellowship (NA07NOS4200018), and by an EFFRD award from Cleveland State University. Numerous students were instrumental in different aspects of the surveys: Mark Lyons, Trevor Prescott, Erin Steiner, Cal Borden, Louie Rundo, and John Hook. Specimens were collected under Ohio Scientific Collecting Permits 194 (2006), 141 (2007), and 11-101 (2008). The Old Woman Creek National Estuarine Research Reserve in Ohio is part of the National Estuarine Research Reserve System (NERRS), established by section 315 of the Coastal Zone Management Act, as amended. Additional information on these preserves and programs is available from the Estuarine Reserves Division, Office for Coastal Management, National Oceanic and Atmospheric Administration, U. S. Department of Commerce, 1305 East West Highway, Silver Spring, MD 20910. -

Fossil Flora and Fauna of Bosnia and Herzegovina D Ela
FOSSIL FLORA AND FAUNA OF BOSNIA AND HERZEGOVINA D ELA Odjeljenje tehničkih nauka Knjiga 10/1 FOSILNA FLORA I FAUNA BOSNE I HERCEGOVINE Ivan Soklić DOI: 10.5644/D2019.89 MONOGRAPHS VOLUME LXXXIX Department of Technical Sciences Volume 10/1 FOSSIL FLORA AND FAUNA OF BOSNIA AND HERZEGOVINA Ivan Soklić Ivan Soklić – Fossil Flora and Fauna of Bosnia and Herzegovina Original title: Fosilna flora i fauna Bosne i Hercegovine, Sarajevo, Akademija nauka i umjetnosti Bosne i Hercegovine, 2001. Publisher Academy of Sciences and Arts of Bosnia and Herzegovina For the Publisher Academician Miloš Trifković Reviewers Dragoljub B. Đorđević Ivan Markešić Editor Enver Mandžić Translation Amra Gadžo Proofreading Amra Gadžo Correction Sabina Vejzagić DTP Zoran Buletić Print Dobra knjiga Sarajevo Circulation 200 Sarajevo 2019 CIP - Katalogizacija u publikaciji Nacionalna i univerzitetska biblioteka Bosne i Hercegovine, Sarajevo 57.07(497.6) SOKLIĆ, Ivan Fossil flora and fauna of Bosnia and Herzegovina / Ivan Soklić ; [translation Amra Gadžo]. - Sarajevo : Academy of Sciences and Arts of Bosnia and Herzegovina = Akademija nauka i umjetnosti Bosne i Hercegovine, 2019. - 861 str. : ilustr. ; 25 cm. - (Monographs / Academy of Sciences and Arts of Bosnia and Herzegovina ; vol. 89. Department of Technical Sciences ; vol. 10/1) Prijevod djela: Fosilna flora i fauna Bosne i Hercegovine. - Na spor. nasl. str.: Fosilna flora i fauna Bosne i Hercegovine. - Bibliografija: str. 711-740. - Registri. ISBN 9958-501-11-2 COBISS/BIH-ID 8839174 CONTENTS FOREWORD ........................................................................................................... -

Proceedings of the Academy of Natural Sciences of Philadelphia
32 PROCEEDINGS OF THE ACADEMY OF [1887. ON NEW GENEEIC FOKMS OF CRETACEOUS MOLLUSCA AND THEIE RELATION TO OTHER FORMS. BY CHARLES A. WHITE. Published by permission of the Director of the United States Geological Survey. The type species of the three generic forms which are described in this article ^ belong to the collections of Cretaceous fossils from Texas, which I am now preparing for publication in one of the memoirs of the U. S. Geological Survey. In their generic charac- teristics all three of them appear to be respectively identical with certain forms which have long been known, but which have been referred to other genera by different authors. The features which I now present as having generic value seem to have been overlooked by those authors, or, so far as they were observed, they were treated as specific characters. Two of these forms belong to the section Melininse of the family Aviculidse. The other is referred to the Crassatellidse, but it departs considerably from the typical section of that family. CRASSATELLIDJE. Genus STEARNSIA (gen. nov.). Shell compressed, subtrihedral or subcircular in marginal out- line; beaks small, closely approximate, prominent by reason of the abrupt sloping away of both the antero-and postero-dorsal borders; lunule and escutcheon both well defined and flattened or excavated; hinge strong, consisting of both cardinal and lateral teeth; cardinal teeth two in the left valve and three in the right; both posterior and anterior lateral teeth long and slender; posterior laterals two in the right valve and one in the left; anterior laterals two in the left valve and one in the right. -

Missouri's Freshwater Mussels
Missouri mussel invaders Two exotic freshwater mussels, the Asian clam (Corbicula and can reproduce at a much faster rate than native mussels. MISSOURI’S fluminea) and the zebra mussel (Dreissena polymorpha), have Zebra mussels attach to any solid surface, including industrial found their way to Missouri. The Asian clam was introduced pipes, native mussels and snails and other zebra mussels. They into the western U.S. from Asia in the 1930s and quickly spread form dense clumps that suffocate and kill native mussels by eastward. Since 1968 it has spread rapidly throughout Missouri restricting feeding, breathing and other life functions. Freshwater and is most abundant in streams south of the Missouri River. In You can help stop the spread of these mussels by not moving the mid-1980s, zebra mussels hitched a ride in the ballast waters bait or boat well water from one stream to another; dump and of freighter ships traveling from Asia to the Great Lakes. They drain on the ground before leaving. Check all surfaces of your have rapidly moved into the Mississippi River basin and boat and trailer for zebra mussels and destroy them, along with westward to Oklahoma. vegetation caught on the boat or trailer. Wash with hot (104˚F) Asian clam and zebra mussel larvae have an advantage here water at a carwash and allow all surfaces to dry in the sun for at because they don’t require a fish host to reach a juvenile stage least five days before boating again. MusselsMusselsSue Bruenderman, Janet Sternburg and Chris Barnhart Zebra mussels attached to a native mussel JIM RATHERT ZEBRA CHRIS BARNHART ASIAN CLAM MUSSEL Shells are very common statewide in rivers, ponds and reservoirs A female can produce more than a million larvae at one time, and are often found on banks and gravel bars.