Essential Roles of Inhibin Beta a in Mouse Epididymal Coiling
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Structure of Pronephros and Development of Mesonephric Kidney in Larvae of Russian Sturgeon, Acipenser Gueldenstaedtii Brandt (Acipenseridae)
Zoologica5 PRONEPHROS Poloniae-AND (2012)-MESONEPHRIC 57/1-4: 5-20-KIDNEY-IN-LARVAE-OF-A.-GUELDENSTAEDTII 5 DOI: 10.2478/v10049-012-0001-6 STRUCTURE OF PRONEPHROS AND DEVELOPMENT OF MESONEPHRIC KIDNEY IN LARVAE OF RUSSIAN STURGEON, ACIPENSER GUELDENSTAEDTII BRANDT (ACIPENSERIDAE) L.S. KRAYUSHKINA*1, A.A. GERASIMOV1, A.A. KIRSANOV1, M.V. MOSYAGINA1, A. OGORZA£EK2 1Department of Ichthyology and Hydrobiology, St. Petersburg State University, 16-th Line 29, 199178, St. Petersburg, Russia, [email protected] 2 Department of Animal Developmental Biology, Zoological Institute, University of Wroclaw, Sienkiewicza 21, 50-335 Wroclaw, Poland. *Corresponding author Abstract. The structure of the pronephros and development of mesonephric kidney in Russian sturgeon larvae, Acipenser gueldenstaedtii Brandt at different stages of early postembryonic development (from hatching to 14 days), were studied with histological and electronic microscopy methods. The larval pronephros is represented by the system of bilaterally located pronephric tubules with ciliated nephrostomes and funnels and exog- enous single glomus, which is not integrated directly into pronephric tubules and located in the pronephric chamber. The glomus is positioned below the dorsal aorta and vascular- ized by its capillaries. The glomus has the same features of the thin structure that are typical of and necessary for the function of a filtering organ. The structure of the prone- phros in acipenserids is discussed and compared with teleosts and amphibians. Histogen- esis of the mesonephric kidney is observed during the period of pronephros functioning; it is complete by the time the larvae transfer to exogenous feeding. At this moment, the pronephros undergoes significant structural degradation. -

Secreted Molecules in Metanephric Induction
J Am Soc Nephrol 11: S116–S119, 2000 Secreted Molecules in Metanephric Induction THOMAS J. CARROLL and ANDREW P. McMAHON Department of Molecular and Cellular Biology, Biological Laboratories, Harvard University, Cambridge, Massachusetts. Abstract. Nearly 50 yr ago, Clifford Grobstein made the ob- the classic model of metanephric induction. The studies of the servation that the ureteric bud induced the nephrogenic mes- classic ureteric inducer performed to date have most likely enchyme to undergo tubulogenesis. Since that discovery, sci- been characterizations of a mesenchyme-specific inducer, entists have attempted to characterize the molecular nature of Wnt-4, and its role in tubulogenesis. Ureteric induction most the inducer. To date, no single molecule that is both necessary likely involves a series of distinct events that provide prolif- and sufficient for nephric induction has been identified. Be- erative, survival, and condensation signals to the mesenchyme, cause of recent insights regarding the role of several secreted integrating the growth of the ureteric system with tubulogen- molecules in tubulogenesis, it has become necessary to revise esis. The developmental biologic processes of the kidney have been logenesis. The conclusion drawn from this discovery was that the subject of intense study for more than 100 yr (for review, the ureteric bud induces tubulogenesis within the surrounding see reference (1). All three vertebrate kidney types (pro- mesenchyme. During further investigation, it was discovered nephros, mesonephros, and metanephros) are derivatives of a that a number of tissues, including, most notably, a dorsal region of the embryo known as the intermediate mesoderm. In portion of the embryonic spinal cord, are able to substitute for mice, a portion of the mesonephric duct, known as the meta- the ureter in this inductive interaction. -

Urinary System Intermediate Mesoderm
Urinary System Intermediate mesoderm lateral mesoderm: somite ectoderm neural NOTE: Intermediate mesoderm splanchnic groove somatic is situated between somites and lateral mesoderm (somatic and splanchnic mesoderm bordering the coelom). All mesoderm is derived from the primary mesen- intermediate mesoderm endoderm chyme that migrated through the notochord coelom (becomes urogenital ridge) primitive streak. Intermediate mesoderm (plus adjacent mesothelium lining the coelom) forms a urogenital ridge, which consists of a laterally-positioned nephrogenic cord (that forms kidneys & ureter) and a medially-positioned gonadal ridge (for ovary/testis & female/male genital tract formation). Thus urinary & genital systems have a common embryonic origin; also, they share common ducts. NOTE: Urine production essentially requires an increased capillary surface area (glomeruli), epithelial tubules to collect plasma filtrate and extract desirable constituents, and a duct system to convey urine away from the body. Kidneys Bilateraly, three kid- mesonephric duct neys develop from the neph- metanephros pronephros rogenic cord. They develop mesonephric tubules chronologically in cranial- mesonephros caudal sequence, and are designated pro—, meso—, Nephrogenic Cord (left) and meta—, respectively. cloaca The pronephros and mesonephros develop similarly: the nephrogenic cord undergoes seg- mentation, segments become tubules, tubules drain into a duct & eventually tubules disintegrate. spinal ganglion 1] Pronephros—consists of (7-8) primitive tubules and a pronephric duct that grows caudally and terminates in the cloaca. The tubules soon degenerate, but the pronephric duct persists as the neural tube mesonephric duct. (The pronephros is not functional, somite except in sheep.) notochord mesonephric NOTE tubule The mesonephros is the functional kidney for fish and am- aorta phibians. The metanephros is the functional kidney body of reptiles, birds, & mammals. -

The Developing Renal, Reproductive, and Respiratory Systems of the African Elephant Suggest an Aquatic Ancestry
Proc. Natl. Acad. Sci. USA Vol. 96, pp. 5555–5558, May 1999 Developmental Biology, Evolution The developing renal, reproductive, and respiratory systems of the African elephant suggest an aquatic ancestry A. P. GAETH*†,R.V.SHORT‡, AND M. B. RENFREE* *Department of Zoology, University of Melbourne, Parkville, Victoria 3052, Australia; and ‡Department of Perinatal Medicine, University of Melbourne, The Royal Women’s Hospital, Carlton, Victoria 3053, Australia Communicated by Judith Kimble, University of Wisconsin, Madison, WI, March 18, 1999 (received for review December 28, 1998) ABSTRACT The early embryology of the elephant has never of the testes, and the precocious development of the trunk could been studied before. We have obtained a rare series of African all have been adaptations to an aquatic lifestyle. elephant (Loxodonta africana) embryos and fetuses ranging in weight from 0.04 to 18.5 g, estimated gestational ages 58–166 MATERIALS AND METHODS days (duration of gestation is '660 days). Nephrostomes, a We have been able to obtain some rare African elephant spec- feature of aquatic vertebrates, were found in the mesonephric imens consisting of one embryo (weight 0.04 g) and six fetuses kidneys at all stages of development whereas they have never (weight ranging from 0.79 to 18.5 g) that were collected from been recorded in the mesonephric kidneys of other viviparous adult females shot in the Kruger National Park, South Africa, mammals. The trunk was well developed even in the earliest between 1993 and 1995, as part of a culling operation to reduce fetus. The testes were intra-abdominal, and there was no evidence elephant numbers in the park. -
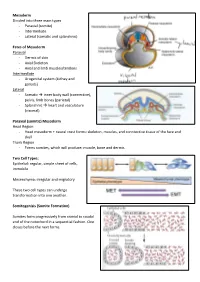
Mesoderm Divided Into Three Main Types - Paraxial (Somite) - Intermediate - Lateral (Somatic and Splanchnic)
Mesoderm Divided into three main types - Paraxial (somite) - Intermediate - Lateral (somatic and splanchnic) Fates of Mesoderm Paraxial - Dermis of skin - Axial Skeleton - Axial and limb muscles/tendons Intermediate - Urogenital system (kidney and gonads) Lateral - Somatic inner body wall (connective), pelvis, limb bones (parietal) - Splanchnic heart and vasculature (visceral) Paraxial (somitic) Mesoderm Head Region - Head mesoderm + neural crest forms: skeleton, muscles, and conntective tissue of the face and skull Trunk Region - Forms somites, which will produce: muscle, bone and dermis Two Cell Types: Epithelial: regular, simple sheet of cells, immobile Mesenchyma: irregular and migratory These two cell types can undergo transformation into one another. Somitogenisis (Somite Formation) Somites form progressively from cranial to caudal end of the notochord in a sequential fashion. One closes before the next forms. Somite Differentiation The somite splits into the epithelial dermamyotome (dermis/muscle) and the messenchymal sclerotome (skeletal). The somite is all paraxial mesoderm. Somite location determines the fate of its associates derma/myo/sclerotomes. Intermediate Mesoderm Urogenital system: - Kidneys - Gonads - Reproductive Duct Systems Runs alongside the paraxial mesoderm. Urogenital System Along mesonephric duct: - Pronephros, mesonephros, and metanephros - Pronephros fall away as gonad develops on ventral-medial side of mesonephros. - Metanephrogenic mesenchyme gives rise to kidney. The mesonephric duct will become the Wolffian duct forming at the nephric bud. The Mullerian duct forms via an invagination on the dorsal side of the nephric duct. The gonad will degenerate one of the two ducts depending on the hormones it produces. XX degenerates Wolffian duct – no testosterone, anti-Mullerian hormone (AMH) not produced, and Mullerian duct can develop in addition to female reproductive organs (ovaries, vagina) XY degenerates Mullerian duct – testosterone, AMH produced, Wolffian duct continues as male reproductive organs (testes, penis) develop. -

Renal Development
RENAL DEVELOPMENT Jon Barasch M.D., Ph.D. Telephone: 305-1890 e-mail: [email protected] SUGGESTED READING: Larsen, 3rd edition, pp 265 (first three paragraphs) - 266, 268-276 and figure 10-10 LEARNING OBJECTIVES: You should be able to: 1. Describe the three kidneys that are produced during development and know what happens to each one. 2. Explain what is meant by ‘reciprocal induction’ and why it poses problems in interpreting experiments in developing kidney. 3. Describe the stages of nephron formation from the renal vesicle. 4. Discuss the regulators of mesenchymal to epithelial transition in the intermediate mesoderm and metanephric mesenchyme and name three molecules mediating conversion. 5. Describe branching morphogenesis and name the three patterns in the developing metanephros. 6. Discuss three key important ligands and their receptors. 7. Discuss the classification of congenital renal abnormalities that are associated with urological abnormalities and the possible underlying mechanisms for their association. SUMMARY: The urogenital system derives from mesenchymal cells by a process of conversion to epithelia. The development of the kidney relies on three mechanisms of epithelial morphogenesis. 1. Some newborn epithelia migrate extensively (Wolffian duct), 2. some undergo branching morphogenesis (ureteric bud) and 3. some produce highly segmented tubules (nephrons). GLOSSARY: Angiotensin II: your favorite vasoconstrictor and regulator of proximal tubule reclamation of NaCl and water by receptor type 1. Receptor type-2 modulates cell growth and seems to play a role in congenital abnormalities. Arcade: a tubule of ureteric bud that induces a few nephrons simultaneously. The nephrons join to a common drainage called a connecting tubule that feeds into the ureteric’s collecting duct. -

Kidney Development and Function in the Fetus
Kidney Development, - and Function in the Fetus Bob Caruthers, CST, PhD The kidneys produce urine, a blood filtrate, and regulate uri- can be seen in the fourth week. The pronephros is complete1 nary volume and composition. These regulatory activities regred by the start of the fifth week. The pronephros form involve balancing water and solute transport, conserving and regresses in a cranial-to-caudal sequence. No pronephric nutrients, eliminating waste products, and regulating acid and glomeruli (cluster of capillaries) have been observed, and no bases. The primary purpose of kidney function is to maintain a vesicles are associated with the pronephric duct. The stable environment in which cellular and tissue metabolic pronephros is, therefore, not active in urine producti0n.Y activity can proceed at an optimal level. The kidneys secrete the hormone renin, erythropoietin and 1.25-dihydroxy vita- MPSONEPHROS min D. Renin helps regulate blood pressure. Erythropoietin The mesonephros originates in the nephrogenic cord that is helps regulate erythrocyte production. 1,25-dihydroxy vitamin part of the intermediate mesoderm. Early mesonephric forma- D plays a role in calcium metabolism. This article will discuss tion is evident before the pronephros has completed its regre: the development of the kidney, and its role in the fetus."' sion. The mesonephros also degenerates in a cranial-to-cauda Since the kidneys are bilateral structures, development sequence. Some of the cranial structures are degenerating in involves both right and left kidneys. During fetal develop- the fifth week of fetal development while the caudal structun ment, three separate nephric structures develop in succession; are still differentiating. -

Organogenesis Part 2 ______
Organogenesis Part 2 ___________________________________ ___________________________________ V. Lateral Plate Mesoderm VI. Endoderm ___________________________________ VII. Development of the Tetrapod Limb VIII. Sex Determination ___________________________________ ___________________________________ ___________________________________ ___________________________________ V. Lateral Plate Mesoderm ___________________________________ ___________________________________ paraxial mesoderm chordamesoderm intermediate mesoderm ___________________________________ ___________________________________ ___________________________________ lateral plate mesoderm ___________________________________ ___________________________________ Lateral Plate Mesoderm ___________________________________ Terminology: - Somatopleure: somatic mesoderm plus ectoderm ___________________________________ - Splanchnopleure: splanchnic mesoderm plus endoderm - Coelom: body cavity forms between them ___________________________________ ___________________________________ ___________________________________ ___________________________________ ___________________________________ Lateral Plate Mesoderm ___________________________________ • The Coelom: ___________________________________ – eventually left and right cavities fuse into one ___________________________________ – runs from neck to anus in vertebrates – portioned off by folds of somatic mesoderm ___________________________________ • pleural cavity: surrounds the thorax and lungs • pericardial cavity: surrounds the -

Odd-Skipped Related 1 Is Required for Development of the Metanephric Kidney and Regulates Formation and Differentiation of Kidne
DEVELOPMENT AND DISEASE RESEARCH ARTICLE 2995 Development 133, 2995-3004 (2006) doi:10.1242/dev.02442 Odd-skipped related 1 is required for development of the metanephric kidney and regulates formation and differentiation of kidney precursor cells Richard G. James1,2,*, Caramai N. Kamei1,2,*, Qingru Wang3, Rulang Jiang3 and Thomas M. Schultheiss1,2,† Formation of kidney tissue requires the generation of kidney precursor cells and their subsequent differentiation into nephrons, the functional filtration unit of the kidney. Here we report that the gene odd-skipped related 1 (Odd1) plays an important role in both these processes. Odd1 is the earliest known marker of the intermediate mesoderm, the precursor to all kidney tissue. It is localized to mesenchymal precursors within the mesonephric and metanephric kidney and is subsequently downregulated upon tubule differentiation. Mice lacking Odd1 do not form metanephric mesenchyme, and do not express several other factors required for metanephric kidney formation, including Eya1, Six2, Pax2, Sall1 and Gdnf. In transient ectopic expression experiments in the chick embryo, Odd1 can promote expression of the mesonephric precursor markers Pax2 and Lim1. Finally, persistent expression of Odd1 in chick mesonephric precursor cells inhibits differentiation of these precursors into kidney tubules. These data indicate that Odd1 plays an important role in establishing kidney precursor cells, and in regulating their differentiation into kidney tubular tissue. KEY WORDS: Kidney, Metanephros, Mesonephros, Chick embryo, Mouse, Osr1, Odd1 INTRODUCTION form a precursor cell population, which both maintains itself at the tips Vertebrate kidney tissue is derived from the intermediate mesoderm of the ureteric bud (via proliferation and/or addition from the (IM), a strip of tissue located adjacent to the somites in the surrounding non-condensed mesenchyme) and gives off cells that developing embryo (Saxen, 1987). -

Development of Urogenital System Dhaka
Mohammad Saiful Islam Phd (Japan) Post doc (Australia) Dept. of Anatomy, Histology & Physiology Sher-e-Bangla Agricultural University Development of Urogenital System Dhaka Intermediate mesoderm forms a urogenital ridge Uro-genital ridge consists of laterally-positioned nephrogenic cord (that becomes kidneys & ureter) and a medially positioned gonadal ridge (for ovary/testis & female/male genital tract formation). Urinary & genital systems have a common embryonic origin (urogenital ridge) and they share common ducts. Kidneys Fig. 1 Development of pro-, meso- and meta-nephros Three kidneys develop chronologically (cranio-caudal) from nephrogenic cord. They are : pronephros, mesonephros and metanephros Development of pronephros and mesonephro: have a similar development Pronephros Consists of (7-8) primitive tubules and a pronephric duct That grows caudally and terminates in the cloaca. The tubules soon degenerate but the pronephric duct persists as the mesonephric duct. Mesonephros Consists of (70-80) tubules induced to form by the mesonephric duct (former pronephric duct) one end of each tubule surrounds a glomerulus (vascular proliferation produced by a branch of the dorsal aorta) The other end of the tubule communicates with the mesonephric duct Eventually, the mesonephros degenerates, but the mesonephric duct becomes epididymis & ductus deferens Metanephros Becomes adult kidney & ureter of mammals, birds, and reptiles Originates in the pelvic region and moves cranially into the abdomen during embryonic differential growth Lobulated initially but becomes smooth in most species. The metanephros originates from two sources: Fig. 2 Development metanephros, ureter a ureteric bud, which grows out of the mesonephric duct near and urinary Bladder the cloaca; the bud develops into the ureter, renal pelvis, and numerous collecting ducts; metanephrogenic mass, which is the caudal region of the nephrogenic cord; the mass forms nephrons. -

Kidney Development
Development of the Urinary System Thomas A. Marino, Ph.D. Department of Anatomy and Cell Biology Temple University School of Medicine Competencies: Upon completion of this section of the course, the student must be able to: ! • Define the three embryonic kidneys. • Know the developmental fate of the pronephros, mesonephros and metanephros. • Understand the origin of cells that develop into the different segments of the kidney tubule. • Compare and contrast the collecting tubule with the nephron. Development of the Kidney • A nephron contains 10,000 cells. • A nephron has at least 12 cell types. • 1 - 3 million collecting tubules are formed. Development of the Kidney • The development of the pronephros • The development of the mesonephros • The development of the metanephros • The development of the kidneys Development of the Pronephros • By day 22 intermediate mesoderm is identified lateral to the paraxial mesoderm. Intermediate Mesoderm Development of the Kidneys • Intermediate mesoderm gives rise to nephrotomes or nephric vesicles in the cervical region. • This is the beginning of the pronephros. • These are non-functional. • They are vestigial remnants that disappear by day 24 or 25. Development of the Kidney Aorta • Glomeruli arise from vessels that branch from the aorta. Development of the Kidneys • By day 23 intermediate mesoderm (orange and navy) is identified lateral to the paraxial mesoderm (red). • Intermediate mesoderm is organized into: • pronephros • mesonephrose • metanephros Development of the Kidneys • Nephric tubules appear in intermediate mesoderm. • Tubules have: – Bowman’s capsule – glomerulus – link to mesonephric duct Development of the Kidneys • As the mesonephros develops, mesonephric tubules form and a mesonephric duct develops. • Mesonephric ducts appear at 24 days. -

Transcriptional Control of Epithelial Differentiation During Kidney Development
J Am Soc Nephrol 14: S9–S15, 2003 Transcriptional Control of Epithelial Differentiation during Kidney Development DAVID RIBES,* EVELYNE FISCHER,† AME´ LIE CALMONT,* and JEROME ROSSERT* *INSERM U489 and The University Pierre and Marie Curie, Paris, France; and †Pasteur Institute, Paris, France. In Mammals, kidney development proceeds in three stages As for many other structures, the combinatorial action of (reviewed in 1). The first two stages lead to the formation of different cell-specific transcription factors is very likely to play transient structures, the pronephros and the mesonephros, and a critical role in the development of the ureteric bud and of the the third stage gives rise to the metanephros, which is the metanephric mesenchyme. In this review, we focus on tran- permanent kidney. However, similar pathways seem to be scription factors that have been shown to play a role in vivo in involved in the development of all three structures. First, the the differentiation of metanephric mesenchymal cells into ep- pronephric tubules and the pronephric duct form at the cervical ithelial or stromal cells and in the differentiation of ureteric bud end of the intermediate mesoderm and fuse to form the prone- cells into epithelial cells of the excretory system. phros. These two components of the pronephros are epithelial structures that arise from an epithelial transformation of neph- Early Differentiation of Metanephric rogenic mesoderm. Second, the Wolffian duct, which is an Mesenchymal Cells extension of the pronephric duct, grows caudally, reaches the In Drosophila melanogaster, the genes eyeless, sine oculis, mesonephric mesenchyme, and induces the formation of the eyes absent, and dachshund form a regulatory network that mesonephros.