N-Alkane Profile of Argemone Mexicana Leaves
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Catalytic Pyrolysis of Plastic Wastes for the Production of Liquid Fuels for Engines
Electronic Supplementary Material (ESI) for RSC Advances. This journal is © The Royal Society of Chemistry 2019 Supporting information for: Catalytic pyrolysis of plastic wastes for the production of liquid fuels for engines Supattra Budsaereechaia, Andrew J. Huntb and Yuvarat Ngernyen*a aDepartment of Chemical Engineering, Faculty of Engineering, Khon Kaen University, Khon Kaen, 40002, Thailand. E-mail:[email protected] bMaterials Chemistry Research Center, Department of Chemistry and Center of Excellence for Innovation in Chemistry, Faculty of Science, Khon Kaen University, Khon Kaen, 40002, Thailand Fig. S1 The process for pelletization of catalyst PS PS+bentonite PP ) t e PP+bentonite s f f o % ( LDPE e c n a t t LDPE+bentonite s i m s n HDPE a r T HDPE+bentonite Gasohol 91 Diesel 4000 3500 3000 2500 2000 1500 1000 500 Wavenumber (cm-1) Fig. S2 FTIR spectra of oil from pyrolysis of plastic waste type. Table S1 Compounds in oils (%Area) from the pyrolysis of plastic wastes as detected by GCMS analysis PS PP LDPE HDPE Gasohol 91 Diesel Compound NC C Compound NC C Compound NC C Compound NC C 1- 0 0.15 Pentane 1.13 1.29 n-Hexane 0.71 0.73 n-Hexane 0.65 0.64 Butane, 2- Octane : 0.32 Tetradecene methyl- : 2.60 Toluene 7.93 7.56 Cyclohexane 2.28 2.51 1-Hexene 1.05 1.10 1-Hexene 1.15 1.16 Pentane : 1.95 Nonane : 0.83 Ethylbenzen 15.07 11.29 Heptane, 4- 1.81 1.68 Heptane 1.26 1.35 Heptane 1.22 1.23 Butane, 2,2- Decane : 1.34 e methyl- dimethyl- : 0.47 1-Tridecene 0 0.14 2,2-Dimethyl- 0.63 0 1-Heptene 1.37 1.46 1-Heptene 1.32 1.35 Pentane, -

Alteration of Volatile Chemical Composition in Tobacco Plants Due to Green Peach Aphid (Myzus Persicae Sulzer) (Hemiptera: Aphididae) Feeding
Song et al.: Green peach aphid (Myzus persicae Sulzer) (Hemiptera: Aphididae) feeding alters volatile composition in tobacco plants - 159 - ALTERATION OF VOLATILE CHEMICAL COMPOSITION IN TOBACCO PLANTS DUE TO GREEN PEACH APHID (MYZUS PERSICAE SULZER) (HEMIPTERA: APHIDIDAE) FEEDING SONG, Y. Z.1,2,3 – GUO, Y. Q.1,2 – CAI, P. M.2,3 – CHEN, W. B.1,2 – LIU, C. M.1,2* 1Biological Control Research Institute, College of Plant Protection, Fujian Agriculture and Forestry University, Fuzhou 350002, China e-mail: [email protected] (Song, Y. Z.); [email protected] (Guo, Y. Q.); [email protected] (Chen, W. B.) 2State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops, Fuzhou 350002, China e-mail: [email protected] (Cai, P. M.) 3Department of Horticulture, College of Tea and Food Science, Wuyi University, Wuyishan 354300, China *Corresponding author e-mail: [email protected]; phone: +86-0591-8378-9420; fax: +86-0591-8378-9421 (Received 10th Jul 2020; accepted 6th Oct 2020) Abstract. In response to insect pest herbivory, plants can generate volatile components that may serve multiple roles as communication signals and defence agents in a multitrophic context. In the present study, the volatile profiles of tobacco plants Nicotiana tabacum L., with and without infestation by sap-sucking aphids Myzus persicae Sulzer, were measured by gas chromatography-mass spectrometry (GC-MS). The results revealed that a total of 10 compounds were identified from healthy tobacco plants, and a total of 14 and 16 compounds were isolated from aphid-infested tobacco plants at 24 and 48 hours after infestation, respectively. -

Synthetic Turf Scientific Advisory Panel Meeting Materials
California Environmental Protection Agency Office of Environmental Health Hazard Assessment Synthetic Turf Study Synthetic Turf Scientific Advisory Panel Meeting May 31, 2019 MEETING MATERIALS THIS PAGE LEFT BLANK INTENTIONALLY Office of Environmental Health Hazard Assessment California Environmental Protection Agency Agenda Synthetic Turf Scientific Advisory Panel Meeting May 31, 2019, 9:30 a.m. – 4:00 p.m. 1001 I Street, CalEPA Headquarters Building, Sacramento Byron Sher Auditorium The agenda for this meeting is given below. The order of items on the agenda is provided for general reference only. The order in which items are taken up by the Panel is subject to change. 1. Welcome and Opening Remarks 2. Synthetic Turf and Playground Studies Overview 4. Synthetic Turf Field Exposure Model Exposure Equations Exposure Parameters 3. Non-Targeted Chemical Analysis Volatile Organics on Synthetic Turf Fields Non-Polar Organics Constituents in Crumb Rubber Polar Organic Constituents in Crumb Rubber 5. Public Comments: For members of the public attending in-person: Comments will be limited to three minutes per commenter. For members of the public attending via the internet: Comments may be sent via email to [email protected]. Email comments will be read aloud, up to three minutes each, by staff of OEHHA during the public comment period, as time allows. 6. Further Panel Discussion and Closing Remarks 7. Wrap Up and Adjournment Agenda Synthetic Turf Advisory Panel Meeting May 31, 2019 THIS PAGE LEFT BLANK INTENTIONALLY Office of Environmental Health Hazard Assessment California Environmental Protection Agency DRAFT for Discussion at May 2019 SAP Meeting. Table of Contents Synthetic Turf and Playground Studies Overview May 2019 Update ..... -

Sampling Dynamics for Volatile Organic Compounds Using Headspace Solid-Phase Microextraction Arrow for Microbiological Samples
separations Communication Sampling Dynamics for Volatile Organic Compounds Using Headspace Solid-Phase Microextraction Arrow for Microbiological Samples Kevin E. Eckert 1, David O. Carter 2 and Katelynn A. Perrault 1,* 1 Laboratory of Forensic and Bioanalytical Chemistry, Forensic Sciences Unit, Division of Natural Sciences and Mathematics, Chaminade University of Honolulu, 3140 Waialae Ave., Honolulu, HI 96816, USA; [email protected] 2 Laboratory of Forensic Taphonomy, Forensic Sciences Unit, Division of Natural Sciences and Mathematics, Chaminade University of Honolulu, 3140 Waialae Avenue, Honolulu, HI 96816, USA; [email protected] * Correspondence: [email protected] Received: 10 August 2018; Accepted: 27 August 2018; Published: 10 September 2018 Abstract: Volatile organic compounds (VOCs) are monitored in numerous fields using several commercially-available sampling options. Sorbent-based sampling techniques, such as solid-phase microextraction (SPME), provide pre-concentration and focusing of VOCs prior to gas chromatography–mass spectrometry (GC–MS) analysis. This study investigated the dynamics of SPME Arrow, which exhibits an increased sorbent phase volume and improved durability compared to traditional SPME fibers. A volatile reference mixture (VRM) and saturated alkanes mix (SAM) were used to investigate optimal parameters for microbiological VOC profiling in combination with GC–MS analysis. Fiber type, extraction time, desorption time, carryover, and reproducibility were characterized, in addition to a comparison with traditional SPME fibers. The developed method was then applied to longitudinal monitoring of Bacillus subtilis cultures, which represents a ubiquitous microbe in medical, forensic, and agricultural applications. The carbon wide range/polydimethylsiloxane (CWR/PDMS) fiber was found to be optimal for the range of expected VOCs in microbiological profiling, and a statistically significant increase in the majority of VOCs monitored was observed. -

Vapor Pressures and Vaporization Enthalpies of the N-Alkanes from C31 to C38 at T ) 298.15 K by Correlation Gas Chromatography
BATCH: je3a29 USER: ckt69 DIV: @xyv04/data1/CLS_pj/GRP_je/JOB_i03/DIV_je030236t DATE: March 15, 2004 1 Vapor Pressures and Vaporization Enthalpies of the n-Alkanes from 2 C31 to C38 at T ) 298.15 K by Correlation Gas Chromatography 3 James S. Chickos* and William Hanshaw 4 Department of Chemistry and Biochemistry, University of MissourisSt. Louis, St. Louis, Missouri 63121, USA 5 6 The temperature dependence of gas-chromatographic retention times for henitriacontane to octatriacontane 7 is reported. These data are used in combination with other literature values to evaluate the vaporization 8 enthalpies and vapor pressures of these n-alkanes from T ) 298.15 to 575 K. The vapor pressure and 9 vaporization enthalpy results obtained are compared with existing literature data where possible and 10 found to be internally consistent. The vaporization enthalpies and vapor pressures obtained for the 11 n-alkanes are also tested directly and through the use of thermochemical cycles using literature values 12 for several polyaromatic hydrocarbons. Sublimation enthalpies are also calculated for hentriacontane to 13 octatriacontane by combining vaporization enthalpies with temperature adjusted fusion enthalpies. 14 15 Introduction vaporization enthalpies of C21 to C30 to include C31 to C38. 54 The vapor pressures and vaporization enthalpies of these 55 16 Polycyclic aromatic hydrocarbons (PAHs) are important larger n-alkanes have been evaluated using the technique 56 17 environmental contaminants, many of which exhibit very of correlation gas chromatography. This technique relies 57 18 low vapor pressures. In the gas phase, they exist mainly entirely on the use of standards in assessment. Standards 58 19 sorbed to aerosol particles. -

Vapor Pressures and Vaporization Enthalpies of the N-Alkanes from 2 C21 to C30 at T ) 298.15 K by Correlation Gas Chromatography
BATCH: je1a04 USER: jeh69 DIV: @xyv04/data1/CLS_pj/GRP_je/JOB_i01/DIV_je0301747 DATE: October 17, 2003 1 Vapor Pressures and Vaporization Enthalpies of the n-Alkanes from 2 C21 to C30 at T ) 298.15 K by Correlation Gas Chromatography 3 James S. Chickos* and William Hanshaw 4 Department of Chemistry and Biochemistry, University of MissourisSt. Louis, St. Louis, Missouri 63121 5 6 The temperature dependence of gas chromatographic retention times for n-heptadecane to n-triacontane 7 is reported. These data are used to evaluate the vaporization enthalpies of these compounds at T ) 298.15 8 K, and a protocol is described that provides vapor pressures of these n-alkanes from T ) 298.15 to 575 9 K. The vapor pressure and vaporization enthalpy results obtained are compared with existing literature 10 data where possible and found to be internally consistent. Sublimation enthalpies for n-C17 to n-C30 are 11 calculated by combining vaporization enthalpies with fusion enthalpies and are compared when possible 12 to direct measurements. 13 14 Introduction 15 The n-alkanes serve as excellent standards for the 16 measurement of vaporization enthalpies of hydrocarbons.1,2 17 Recently, the vaporization enthalpies of the n-alkanes 18 reported in the literature were examined and experimental 19 values were selected on the basis of how well their 20 vaporization enthalpies correlated with their enthalpies of 21 transfer from solution to the gas phase as measured by gas 22 chromatography.3 A plot of the vaporization enthalpies of 23 the n-alkanes as a function of the number of carbon atoms 24 is given in Figure 1. -
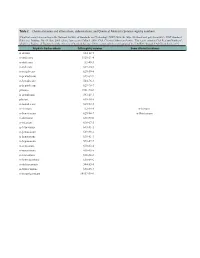
Table 2. Chemical Names and Alternatives, Abbreviations, and Chemical Abstracts Service Registry Numbers
Table 2. Chemical names and alternatives, abbreviations, and Chemical Abstracts Service registry numbers. [Final list compiled according to the National Institute of Standards and Technology (NIST) Web site (http://webbook.nist.gov/chemistry/); NIST Standard Reference Database No. 69, June 2005 release, last accessed May 9, 2008. CAS, Chemical Abstracts Service. This report contains CAS Registry Numbers®, which is a Registered Trademark of the American Chemical Society. CAS recommends the verification of the CASRNs through CAS Client ServicesSM] Aliphatic hydrocarbons CAS registry number Some alternative names n-decane 124-18-5 n-undecane 1120-21-4 n-dodecane 112-40-3 n-tridecane 629-50-5 n-tetradecane 629-59-4 n-pentadecane 629-62-9 n-hexadecane 544-76-3 n-heptadecane 629-78-7 pristane 1921-70-6 n-octadecane 593-45-3 phytane 638-36-8 n-nonadecane 629-92-5 n-eicosane 112-95-8 n-Icosane n-heneicosane 629-94-7 n-Henicosane n-docosane 629-97-0 n-tricosane 638-67-5 n-tetracosane 643-31-1 n-pentacosane 629-99-2 n-hexacosane 630-01-3 n-heptacosane 593-49-7 n-octacosane 630-02-4 n-nonacosane 630-03-5 n-triacontane 638-68-6 n-hentriacontane 630-04-6 n-dotriacontane 544-85-4 n-tritriacontane 630-05-7 n-tetratriacontane 14167-59-0 Table 2. Chemical names and alternatives, abbreviations, and Chemical Abstracts Service registry numbers.—Continued [Final list compiled according to the National Institute of Standards and Technology (NIST) Web site (http://webbook.nist.gov/chemistry/); NIST Standard Reference Database No. -

Global Journal of Science Frontier Research: B Chemistry
Online ISSN : 2249-4626 Print ISSN : 0975-5896 Chemistry and Medicinal Francolin and Turkey Theophylline and Caffeine Synthesis and Characterization VOLUME 13 ISSUE 3 VERSION 1.0 Global Journal of Science Frontier Research: B Chemistry Global Journal of Science Frontier Research: B C hemistry Vol ume 13 Issue 3 (Ver. 1.0) Open Association of Research Society © Global Journal of Science Global Journals Inc. Frontier Research .2013 . (A Delaware USA Incorporation with “Good Standing”; Reg. Number: 0423089) Sponsors: Open Association of Research Society All rights reserved. Open Scientific Standards This is a special issue published in version 1.0 of “Global Journal of Science Frontier Publisher’s Headquarters office Research.” By Global Journals Inc. All articles are open access articles distributed Global Journals Inc., Headquarters Corporate Office, under “Global Journal of Science Frontier Research” Cambridge Office Center, II Canal Park, Floor No. Reading License, which permits restricted use. 5th, Cambridge (Massachusetts), Pin: MA 02141 Entire contents are copyright by of “Global United States Journal of Science Frontier Research” unless USA Toll Free: +001-888-839-7392 otherwise noted on specific articles. USA Toll Free Fax: +001-888-839-7392 No part of this publication may be reproduced or transmitted in any form or by any means, Offset Typesetting electronic or mechanical, including photocopy, recording, or any information Open Association of Research Society, Marsh Road, storage and retrieval system, without written permission. Rainham, Essex, London RM13 8EU United Kingdom. The opinions and statements made in this book are those of the authors concerned. Ultraculture has not verified and neither confirms nor denies any of the foregoing and Packaging & Continental Dispatching no warranty or fitness is implied. -

Comparative Analysis of Volatile Compounds of Gamma-Irradiated Mutants of Rose (Rosa Hybrida)
plants Article Comparative Analysis of Volatile Compounds of Gamma-Irradiated Mutants of Rose (Rosa hybrida) Jaihyunk Ryu 1 , Jae Il Lyu 1 , Dong-Gun Kim 1, Jung-Min Kim 1, Yeong Deuk Jo 1 , Si-Yong Kang 2, Jin-Baek Kim 1 , Joon-Woo Ahn 1 and Sang Hoon Kim 1,* 1 Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute, Jeollabuk-do 56212, Korea; [email protected] (J.R.); [email protected] (J.I.L.); [email protected] (D.-G.K.); [email protected] (J.-M.K.); [email protected] (Y.D.J.); [email protected] (J.-B.K.); [email protected] (J.-W.A.) 2 Department of Horticulture, College of Industrial Sciences, Kongju National University, Yesan 32439, Korea; [email protected] * Correspondence: [email protected]; Tel.: +82-63-570-3318 Received: 22 July 2020; Accepted: 14 September 2020; Published: 17 September 2020 Abstract: Roses are one of the most important floricultural crops, and their essential oils have long been used for cosmetics and aromatherapy. We investigated the volatile compound compositions of 12 flower-color mutant variants and their original cultivars. Twelve rose mutant genotypes were developed by treatment with 70 Gy of 60Co gamma irradiation of six commercial rose cultivars. Essential oils from the flowers of the 18 genotypes were analyzed by gas chromatography–mass spectrometry. Seventy-seven volatile compounds were detected, which were categorized into six classes: Aliphatic hydrocarbons, aliphatic alcohols, aliphatic ester, aromatic compounds, terpene alcohols, and others. Aliphatic (hydrocarbons, alcohols, and esters) compounds were abundant categories in all rose flowers. -

Exhaust Emission Profiles for EPA SPECIATE Database
Exhaust Emission Profiles for EPA SPECIATE Database: Energy Policy Act (EPAct) Low-Level Ethanol Fuel Blends and Tier 2 Light- Duty Vehicles Exhaust Emission Profiles for EPA SPECIATE Database: Energy Policy Act (EPAct) Low-Level Ethanol Fuel Blends and Tier 2 Light- Duty Vehicles Assessment and Standards Division Office of ransportationT and Air Quality U.S. Environmental Protection Agency EPA-420-R-09-002 June 2009 Table of Contents 1.0 Introduction....................................................................................................................... 1 1.1 Background..................................................................................................................... 1 1.2 Purpose of the Project ..................................................................................................... 1 2.0 Methods.............................................................................................................................. 2 2.1 Exhaust Emissions Data.................................................................................................. 2 2.2 Calculation of Composite Profiles.................................................................................. 3 2.3 Adjustment of E0 and E10 profiles with an In-Use Gasoline......................................... 3 2.4 Assignment of SPECIATE Identification Numbers ....................................................... 4 3.0 Results ............................................................................................................................... -

Oral Transfer of Chemical Cues, Growth Proteins and Hormones in Social Insects
Oral transfer of chemical cues, growth proteins and hormones in social insects LeBoeuf, A.C.1,2, Waridel, P.3, Brent, C.S.4, Gonçalves, A.N. 5,8, Menin, L. 6, Ortiz, D. 6, Riba- Grognuz, O. 2, Koto, A.7, Soares Z.G. 5,8, Privman, E. 9, Miska, E.A. 8,10,11, Benton, R.*1 and Keller, L.*2 * order decided on a coin-toss as these authors contributed equally. 1. Center for Integrative Genomics, University of Lausanne, CH-1015 Lausanne, Switzerland 2. Department of Ecology and Evolution, University of Lausanne, CH-1005 Lausanne, Switzerland 3. Protein Analysis Facility, University of Lausanne, 1015 Lausanne, Switzerland. 4. Arid Land Agricultural Research Center, USDA-ARS, 21881 N. Cardon Lane, Maricopa, Arizona, 85138, U.S.A. 5. Department of Biochemistry and Immunology, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Minas Gerais, CEP 30270-901, Brasil. 6. Institute of Chemical Sciences and Engineering, Ecole Polytechnique Fédérale de Lausanne, 1015- Lausanne, Switzerland. 7. The Department of Genetics, Graduate School of Pharmaceutical Sciences, The University of Tokyo. 8. Gurdon Institute, University of Cambridge, Tennis Court Road, Cambridge, CB2 1QN, United Kingdom 9. Department of Evolutionary and Environmental Biology, Institute of Evolution, University of Haifa, Haifa 3498838, Israel.Department of Genetics, University of Cambridge, Downing Street, Cambridge, CB2 3EH, United Kingdom 10. Wellcome Trust Sanger Institute, Wellcome Trust Genome Campus, Cambridge, CB10 1SA, United Kingdom Correspondence to: Adria C. LeBoeuf ([email protected]), Richard Benton ([email protected]) or Laurent Keller ([email protected]) 1 1 Abstract 2 3 Social insects frequently engage in oral fluid exchange – trophallaxis – between adults, and 4 between adults and larvae. -

UST and ASTM Petrochemical Standards
UST and ASTM Petrochemical Standards Your essential resource for Agilent ULTRA chemical standards Table of contents Introduction 3 Pennsylvania – GRO and PAH 22 About Agilent standards 3 Tennessee – GRO and DRO 22 Products 3 Texas – TNRCC Method 1005, 1006 23 Markets 3 Washington – Volatile Petroleum Hydrocarbons (VPH) method 24 Custom products 3 Washington – Extractable Petroleum Hydrocarbons Quality control laboratory 4 (EPH) method 25 Quality control validation levels 4 Washington and Oregon – Total Petroleum Hydrocarbons Triple certification 5 (NWTPH) methods 26 Level 2 reference material Certificate of Analysis 6 Maine – GRO and DRO 26 GHS compliance 7 Internal and surrogate standards for Underground Storage Tank (UST) testing 27 Underground Storage Tank (UST) Standards 8 EPA Method 1664A 28 Alaska – Method AK 101, AK 102, AK 103 10 EPA Method 418.1 28 Arizona – Method 8015AZ 11 Hydrocarbon fuel standards 29 California – PVOC and WIP 12 Weathered hydrocarbon fuel standards 30 Connecticut – ETPH method 12 EN 14105:2003 31 Florida – Method FL-PRO 13 Iowa – Methods OA-1, OA-2 14 ASTM Methods 32 Kansas – TPH method 15 ASTM Method D6584 32 Kansas modified 8015 (LRH) 15 ASTM Method D1387 33 Maine – Method 4.1.25, 4.2.17 16 ASTM Method D3710 34 Massachusetts – Volatile Petroleum Hydrocarbons ASTM Method D4815 34 (VPH) method 17 ASTM Method D5453 35/36 Massachusetts – Extractable Petroleum Hydrocarbons ASTM Methods D3120, D3246, D3961 36 (EPH) method 18 ASTM Method D4629 37 Shooters – Open and shoot spiking standards 19 ASTM Method D5762 38 Michigan – GRO and PNA 20 ASTM Method D4929 38 Mississippi – GRO, DRO, and PAH 20 ASTM Method D5808 38 New Jersey – OQA-QAM-025 21 New York – STARS compounds 22 Agilent Service and Support 39 2 UST and ASTM Petrochemical Standards Introduction About Agilent standards Agilent is a global leader in chromatography and spectroscopy, as well as an expert in chemical standards manufacturing.