Protected Triazabutadienes in Chemical Biology
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

June 2009 MONTHLY MEETING : PLEASE PAY YOUR DUES: WHEN? Monday, June 22, 2009 – the Fourth Monday of in Accordance with the By-Laws, Dues, in the the Month
Spore-Addct The Newsletter of the Pikes PeakTime Mycological Society 1974 – 2009 Vol. XXXV ISSUE 3 June 2009 MONTHLY MEETING : PLEASE PAY YOUR DUES: WHEN? Monday, June 22, 2009 – The fourth Monday of In accordance with the By-laws, dues, in the the month. amount of $15.00, are due and payable on or before the April monthly meeting. If you still WHAT TIME? 6:30 pm; the meeting will come to order at have not paid, please pay at the June meeting or 7:00 pm. mail the payment to PPMS, PO Box 39, Colorado Springs, CO 80901-0039. Thanks! WHERE? Pikes Peak National Bank, 2401 W. Colorado Ave. (across from Bancroft Park). Enter at the door on Colorado Ave., just west of the bank door. There you will find stairs and an elevator. You may use either. The FORAY REPORT: room is on the second floor near the head of the stairs. May 30th 2009: Ten of us (including 2 newcomers to WEBSITE: http://www.pikespeakmushrooms.com/ PPMS) gathered at the Red Rocks Safeway parking lot at 8:30 a.m. All had either been contacted by e-mail or PROGRAM: personal phone call. All club forays will be scheduled like this. Our eager forayers found only 6 small morels, A NAMA Educational Program: 2 L.B.Ms, and some dubious polypors. We did come “Guide to the Major Genera of Gilled Mushrooms: The across 6 Calypso orchids (unaccompanied by morels). Light Spored Mushrooms I: Pluteaceae, Pleurotaceae, It was a beautiful day on Rampart Range Road. Rain Entolomataceae, Marasmiaceae and Others” and warmer temperatures have brought out city This program covers the best edible and most mushrooms galore. -

Screening of Potentially Genotoxic Impurities in Pharmaceuticals by LC-MS and CE-MS
Screening of potentially genotoxic impurities in pharmaceuticals by LC-MS and CE-MS Screening van potentieel genotoxische onzuiverheden in farmaceutische producten met LC-MS en CE-MS (met een samenvatting in het Nederlands) Proefschrift ter verkrijging van de graad van doctor aan de Universiteit Utrecht op gezag van de rector magnificus, prof. dr. G.J. van der Zwaan, ingevolge van het besluit van het college voor promoties in het openbaar te verdedigen op maandag 21 november 2016 des middags te 2.30 uur. door Anthonius Maria van Wijk geboren op 3 juli 1968 te Oostflakkee Promotor: Prof. dr. G.J. de Jong Copromotor: Dr. H.A.G. Niederländer Index Chapter 1 Introduction 5 Chapter 2 The potential for screening of potentially genotoxic impurities 19 in pharmaceutical products Chapter 3 A new approach for generic screening and quantitation of 71 potential genotoxic alkylation compounds by pre-column derivatization and LC-MS/MS analysis Chapter 4 A new derivatization reagent for LC–MS/MS screening of 91 potential genotoxic alkylation compounds Chapter 5 Capillary electrophoresis – mass spectrometry for impurity 115 profiling of basic pharmaceuticals using non-volatile background electrolytes Chapter 6 Sensitive CE-MS analysis of potentially genotoxic alkylation 137 compounds using derivatization and electrokinetic injection Chapter 7 Summary and general conclusions 159 Summary in Dutch/Nederlandse samenvatting 174 Curriculum Vitae 182 List of publications 183 Acknowledgements/Dankwoord 184 Chapter 1 Introduction 5 Chapter 1 1.1 Control of genotoxic impurities Trace analysis of potential genotoxic impurities (PGIs) is one of the greatest challenges in pharmaceutical analysis. Traces of toxic impurities, i.e. -

Synthetic and Naturally Occurring Hydrazines As Possible Cancer Causative Agents
[CANCER RESEARCH 35, 3693-3697 December 1975] Synthetic and Naturally Occurring Hydrazines as Possible Cancer Causative Agents Bela Toth' The Eppley Institute for Research in Cancer, University of Nebraska Medical Center, Omaha, Nebraska 68105 SUMMARY SYNTHETIC HYDRAZINES The various synthetic substituted hydrazines, which cause tumors in animals, are briefly enumerated. To date, 19 of Studies on the carcinogenic potentialities of synthetic them have proved to be tumorigenic in animals. A number substituted hydrazines began in 1962, when it was shown of these chemicals are found today in the environment, in that the base compound hydrazine sulfate induced lung industry, in agriculture, and in medicine, and the human neoplasms in mice (1). Subsequently, a series of hydrazine population is exposed to a certain degree to some of them. derivatives were investigated in various laboratories for Hydrazine also occurs in nature in tobacco and tobacco tumor-inducing capabilities. These studies clearly demon smoke. The three other naturally occurring hydrazine strated that these chemicals are indeed powerful tumori compounds are N-methyl-N-formylhydrazine, which oc genic substances in mice, hamsters, and rats, due to their curs in the wild edible mushroom, Gyromitra esculenta, tumor-inducing abilities in the intestines, brain, lungs, and @-N-[―y-L(+)-glutamylJ-4-hydroxymethylphenyl blood vessels, liver, breasts, kidneys, etc. Now, we know of hydrazine and 4-hydroxymethylphenylhydrazine, whkh 19 hydrazine derivatives that have been shown to be tumor are found in the commonly eaten cultivated mushroom, inducers. These include, in addition to hydrazine (1, 32), Agaricus bisporus. Tumorigenesis studies with the natu methyl- (35, 40), 1,2-dimethyl- (6, 27, 36, 46, 52), 1,1- rally occurring hydrazines are in progress. -

Hydrazine 1 Hydrazine
Hydrazine 1 Hydrazine Hydrazine Identifiers [1] CAS number 302-01-2 , 7803-57-8 (hydrate) [2] EC number 206-114-9 UN number 2029 (anhydrous) 2030 (aq. soln., 37–64%) 3293 (aq. soln., <37%) RTECS number MU7175000 Properties Molecular formula N H 2 4 Molar mass 32.05 g/mol (anhydrous) 50.06 g/mol (hydrate) Appearance Colourless liquid Density 1.0045 g/cm3 (anhydrous) 1.032 g/cm3 (hydrate) Melting point 1 °C (274 K, anhydrous) -51.7 °C (hydrate) Boiling point 114 °C (387 K), anhydrous 119 °C (hydrate) Solubility in water miscible Acidity (pK ) 8.1 a Refractive index (n ) [3] D 1.46044 (22 °C, anhydrous) 1.4284 (hydrate) Viscosity 0.876 cP (25 °C) Structure Hydrazine 2 Molecular shape pyramidal at N [4] Dipole moment 1.85 D Hazards [5] MSDS ICSC 0281 EU Index 007-008-00-3 EU classification Carc. Cat. 2 Toxic (T) Corrosive (C) Dangerous for the environment (N) R-phrases R45, R10, R23/24/25, R34, R43, R50/53 S-phrases S53, S45, S60, S61 NFPA 704 Flash point 52 °C Autoignition 24–270 °C (see text) temperature Explosive limits 1.8–100% LD [6] 50 59–60 mg/kg (oral in rats, mice) Related compounds Related nitrogen hydrides Ammonia Hydrazoic acid Related compounds monomethylhydrazine dimethylhydrazine phenylhydrazine [7] (what is this?) (verify) Except where noted otherwise, data are given for materials in their standard state (at 25 °C, 100 kPa) Infobox references Hydrazine is an inorganic chemical compound with the formula N H . It is a colourless liquid with an 2 4 ammonia-like odor and is derived from the same industrial chemistry processes that manufacture ammonia. -

SOMA NEWS EDITOR Tom Cruckshank [email protected] Meet at Woodside Campground in Salt Point State Park at 10 AM
VOLUME 22 ISSUE 1 September 2009 SOMA IS AN EDUCATIONAL ORGANIZATION DEDICATED TO MYCOLOGY. WE ENCOURAGE ENVIRONMENTAL AWARENESS BY SHARING OUR ENTHUSIASM THROUGH PUBLIC PARTICIPATION AND GUIDED FORAYS. 2009/2010 SPEAKER OF THE MONTH SEASON CALENDAR David Campbell September Sept. 17th » Meeting—7pm September 17th—7pm—Farm Bureau Sonoma County Farm Bureau Speaker: David Campbell Rocky Monsoon Mushrooms: Foray Sept. 19th » Salt Point A peek at summer fungi of southwestern Colorado October avid Campbell… has been collecting, studying, eating, teaching and writing about Oct. 15th » Meeting—7pm D wild mushrooms for 40 years. He has served many years on the council of the Sonoma County Farm Bureau Mycological Society of San Francisco (MSSF), including 2 years as president. For many Speaker: Don Hemmes years, David has served the San Francisco Poison Control Center for mushroom poisoning incident response in Marin County and the greater Bay Area. With a primary Foray Oct. 17th » Salt Point focus on edible and poisonous mushrooms, he leads innumerable fungal forays for November MSSF and the Sonoma Mycological Association (SOMA). David was foray leader and event facilitator for several years at author David Arora's annual mycological field Nov. 20th » Meeting—7pm seminars. As an expert mycophagist (one who safely eats a wide variety of wild Sonoma County Farm Bureau mushrooms) and experienced outdoor group foray leader, David is ‘Foray Director’ for Speaker: TBA Wild About Mushrooms Company, guiding organized wild mushroom adventures, locally Foray Nov. 21st » Salt Point and afar. He recently formed his own company, MycoVentures, Inc., expanding his horizons to include the rest of the planet, for events such as Porcini hunts in the December Potluck on the 11th. -

AROMATIC NUCLEOPHILIC SUBSTITUTION-PART -2 Electrophilic Substitution
Dr. Tripti Gangwar AROMATIC NUCLEOPHILIC SUBSTITUTION-PART -2 Electrophilic substitution ◦ The aromatic ring acts as a nucleophile, and attacks an added electrophile E+ ◦ An electron-deficient carbocation intermediate is formed (the rate- determining step) which is then deprotonated to restore aromaticity ◦ electron-donating groups on the aromatic ring (such as -OH, -OCH3, and alkyl) make the reaction faster, since they help to stabilize the electron-poor carbocation intermediate ◦ Lewis acids can make electrophiles even more electron-poor (reactive), increasing the reaction rate. For example FeBr3 / Br2 allows bromination to occur at a useful rate on benzene, whereas Br2 by itself is slow). In fact, a substitution reaction does occur! (But, as you may suspect, this isn’t an electrophilic aromatic substitution reaction.) In this substitution reaction the C-Cl bond breaks, and a C-O bond forms on the same carbon. The species that attacks the ring is a nucleophile, not an electrophile The aromatic ring is electron-poor (electrophilic), not electron rich (nucleophilic) The “leaving group” is chlorine, not H+ The position where the nucleophile attacks is determined by where the leaving group is, not by electronic and steric factors (i.e. no mix of ortho– and para- products as with electrophilic aromatic substitution). In short, the roles of the aromatic ring and attacking species are reversed! The attacking species (CH3O–) is the nucleophile, and the ring is the electrophile. Since the nucleophile is the attacking species, this type of reaction has come to be known as nucleophilic aromatic substitution. n nucleophilic aromatic substitution (NAS), all the trends you learned in electrophilic aromatic substitution operate, but in reverse. -

The Automated Flow Synthesis of Fluorine Containing Organic Compounds
The automated flow synthesis of fluorine containing organic compounds by CHANTAL SCHOLTZ Submitted in partial fulfilment of the requirements for the degree PHILOSOPHAE DOCTOR In the Faculty of Natural & Agricultural Sciences UNIVERSITY OF PRETORIA PRETORIA Supervisor: Dr D.L. Riley February 2019 DECLARATION I, Chantal Scholtz declare that the thesis/dissertation, which I hereby submit for the degree PhD Chemistry at the University of Pretoria, is my own work and has not previously been submitted by me for a degree at this or any other tertiary institution. Signature :.......................................... Date :..................................... ii ACKNOWLEDGEMENTS I would herewith sincerely like to show my gratitude to the following individuals for their help, guidance and assistance throughout the duration of this project: My supervisor, Doctor Darren Riley, for his knowledge and commitment. Thank you for being a fantastic supervisor and allowing me the opportunity to learn so many valuable skills. My husband, Clinton, for all your love, support and patience and for always being there for me. You are the best. My family, for all the encouragement and support you gave me as well as always believing in me. I will always appreciate what you have done for me. Mr Drikus van der Westhuizen and Mr Johan Postma for their assistance at the Pelchem laboratories with product isolation and characterisation. Dr Mamoalosi Selepe for NMR spectroscopy services, Jeanette Strydom for XRF services and Gerda Ehlers at the UP library for her invaluable assistance. All my friends and colleagues for the continuous moral support, numerous helpful discussions and necessary coffee breaks. My colleagues at Chemical Process Technologies for their ongoing support and motivation, especially Dr Hannes Malan and Prof. -

(10) Patent No.: US 8119385 B2
US008119385B2 (12) United States Patent (10) Patent No.: US 8,119,385 B2 Mathur et al. (45) Date of Patent: Feb. 21, 2012 (54) NUCLEICACIDS AND PROTEINS AND (52) U.S. Cl. ........................................ 435/212:530/350 METHODS FOR MAKING AND USING THEMI (58) Field of Classification Search ........................ None (75) Inventors: Eric J. Mathur, San Diego, CA (US); See application file for complete search history. Cathy Chang, San Diego, CA (US) (56) References Cited (73) Assignee: BP Corporation North America Inc., Houston, TX (US) OTHER PUBLICATIONS c Mount, Bioinformatics, Cold Spring Harbor Press, Cold Spring Har (*) Notice: Subject to any disclaimer, the term of this bor New York, 2001, pp. 382-393.* patent is extended or adjusted under 35 Spencer et al., “Whole-Genome Sequence Variation among Multiple U.S.C. 154(b) by 689 days. Isolates of Pseudomonas aeruginosa” J. Bacteriol. (2003) 185: 1316 1325. (21) Appl. No.: 11/817,403 Database Sequence GenBank Accession No. BZ569932 Dec. 17. 1-1. 2002. (22) PCT Fled: Mar. 3, 2006 Omiecinski et al., “Epoxide Hydrolase-Polymorphism and role in (86). PCT No.: PCT/US2OO6/OOT642 toxicology” Toxicol. Lett. (2000) 1.12: 365-370. S371 (c)(1), * cited by examiner (2), (4) Date: May 7, 2008 Primary Examiner — James Martinell (87) PCT Pub. No.: WO2006/096527 (74) Attorney, Agent, or Firm — Kalim S. Fuzail PCT Pub. Date: Sep. 14, 2006 (57) ABSTRACT (65) Prior Publication Data The invention provides polypeptides, including enzymes, structural proteins and binding proteins, polynucleotides US 201O/OO11456A1 Jan. 14, 2010 encoding these polypeptides, and methods of making and using these polynucleotides and polypeptides. -

S.T.E.T.Women's College, Mannargudi Semester Iii Ii M
S.T.E.T.WOMEN’S COLLEGE, MANNARGUDI SEMESTER III II M.Sc., CHEMISTRY ORGANIC CHEMISTRY - II – P16CH31 UNIT I Aliphatic nucleophilic substitution – mechanisms – SN1, SN2, SNi – ion-pair in SN1 mechanisms – neighbouring group participation, non-classical carbocations – substitutions at allylic and vinylic carbons. Reactivity – effect of structure, nucleophile, leaving group and stereochemical factors – correlation of structure with reactivity – solvent effects – rearrangements involving carbocations – Wagner-Meerwein and dienone-phenol rearrangements. Aromatic nucleophilic substitutions – SN1, SNAr, Benzyne mechanism – reactivity orientation – Ullmann, Sandmeyer and Chichibabin reaction – rearrangements involving nucleophilic substitution – Stevens – Sommelet Hauser and von-Richter rearrangements. NUCLEOPHILIC SUBSTITUTION Mechanism of Aliphatic Nucleophilic Substitution. Aliphatic nucleophilic substitution clearly involves the donation of a lone pair from the nucleophile to the tetrahedral, electrophilic carbon bonded to a halogen. For that reason, it attracts to nucleophile In organic chemistry and inorganic chemistry, nucleophilic substitution is a fundamental class of reactions in which a leaving group(nucleophile) is replaced by an electron rich compound(nucleophile). The whole molecular entity of which the electrophile and the leaving group are part is usually called the substrate. The nucleophile essentially attempts to replace the leaving group as the primary substituent in the reaction itself, as a part of another molecule. The most general form of the reaction may be given as the following: Nuc: + R-LG → R-Nuc + LG: The electron pair (:) from the nucleophile(Nuc) attacks the substrate (R-LG) forming a new 1 bond, while the leaving group (LG) departs with an electron pair. The principal product in this case is R-Nuc. The nucleophile may be electrically neutral or negatively charged, whereas the substrate is typically neutral or positively charged. -

Influence of Storage and Household Processing on the Agaritine Content of the Cultivated Agaricus Mushroom
Food Additives and Contaminants, 2002, Vol. 19, No. 9, 853±862 In¯uence of storage and household processing on the agaritine content of the cultivated Agaricus mushroom V. Schulzova , J. HajsÏ lova *, R. Peroutka , J. (around 10% left after 2 h). Dry baking of the y y y Gry‡ and H. C. Andersson} cultivated mushroom, a process similar to pizza baking, Institute of Chemical Technology, Department of Food Chemistry y reduced the agaritine content by approximately 25%, and Analysis, Technicka 3, 166 28 Prague 6, Czech Republic; whereas frying in oil or butter or deep frying resulted in ‡ Danish Veterinary and Food Administration, Mùrkhùj Bygade 19, DK-2860 Sùborg, Denmark; } National Food Administration, a more marked reduction (35±70%). Microwave pro- Box 622, SE-751 26 Uppsala, Sweden. cessing of the cultivated mushrooms reduced the agar- itine content to one-third of the original level. Thus, the exposure to agaritine was substantially less when (Received 20 June 2001; revised 3 May 2002; accepted 19 consuming processed Agaricus mushrooms as com- May 2002) pared with consuming the raw mushrooms. However, it is not yet known to what extent agaritine and other Agaritine (N-(g-l(+)-glutamyl)-4-hydroxymethyl- phenylhydrazin e derivatives occurring in the cultivated phenylhydrazine ) was identi®ed and quanti®ed by mushroom are degraded into other biologically active high-pressure liquid chromatography and used as a compounds during the cooking procedure. marker for the occurrence of phenylhydrazine deriva- tives in the cultivated Agaricus bitorquis and A. garicus hortensis mushrooms. Although relatively high Keywords : agaritine, Agaricus, Agaricus bitorquis, levels of agaritine (around 700 mg kg±1) could be found Agaricus hortensis, storage, household processing, in freshly harvested A. -

Synthesis of Organobromines As a Tool for Their Characterisation and Environmental Occurrence Assessment
Synthesis of organobromines as a tool for their characterisation and environmental occurrence assessment Andreas Rydén Department of Materials and Environmental Chemistry Stockholm University Stockholm 2013 i Doctoral Thesis 2013 Department of Materials and Environmental Chemistry Stockholm University SE-106 91 Stockholm Sweden Abstract Polybrominated diphenyl ethers (PBDEs) have been intensively used as flame retardants (FRs) and have become ubiquitous environmental pollutants. PBDEs form hydroxylated PBDEs (OH-PBDEs) as metabolites. Further, some OH-PBDEs and methoxy-PBDEs (MeO-PBDEs) are natural products. These are all compounds of environmental and health concern and it is therefore important to confirm their identity and to assess their environmental levels and toxicities. Hence, it is vital to obtain authentic reference standards of individual PBDEs and OH/MeO-PBDEs. The thesis main aim was to develop synthesis methods of congener specific PBDEs, OH- and MeO-PBDEs. The second aim was to identify and quantify PBDEs, OH- and MeO-PBDEs in environmental samples. The third was to propose an abbreviation system for FRs. O-Arylation of brominated phenols, using either symmetrical or unsymmetrical brominated diphenyliodonium salts, was selected for synthesis of PBDEs and OH- /MeO-PBDEs. A total of 16 MeO-PBDEs, 11 OH-PBDEs, 1 diMeO-PBDE and 1 EtO-MeO-PBDE were synthesised. Three novel unsymmetrical diaryliodonium triflates were synthesised and used in synthesis. Optimisations were made to construct a reliable general method for congener specific PBDE synthesis, which was used in the synthesis of 8 representative PBDE congeners. The products were generally characterised by electron ionisation mass spectrometry (EIMS) and nuclear magnetic resonance (NMR) spectroscopy. -
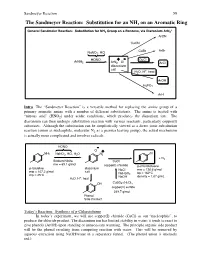
The Sandmeyer Reaction: Substitution for an NH2 on an Aromatic Ring
Sandmeyer Reaction 59 The Sandmeyer Reaction: Substitution for an NH2 on an Aromatic Ring + General Sandmeier Reaction: Substitution for NH2 Group on a Benzene, via Diazonium ArN2 ArCN CuCN CuBr ArBr NaNO2, HCl HONO + - CuCl ArNH2 ArN2 Cl ArCl diazonium salt + H2O, H , heat ArOH H3PO2 ArH Intro The “Sandmeyer Reaction” is a versatile method for replacing the amine group of a primary aromatic amine with a number of different substitutents. The amine is treated with “nitrous acid” (HNO2) under acidic conditions, which produces the diazonium ion. The diazonium can then undergo substitution reaction with various reactants, particularly copper(I) substrates. Although the substitution can be simplistically viewed as a direct ionic substitution reaction (anion as nucleophile, molecular N2 as a premier leaving group), the actual mechanism is actually more complicated and involves radicals. HONO - Cl + NH2 NaNO2, HCl, H2O N2 Cl + N2 Sodium Nitrite CuCl mw = 69.1 g/mol copper(I) chloride p-chlorotoluene p-toluidine diazonium NaCl mw = 126.6 g/mol mw = 107.2 g/mol salt NaHSO bp = 162ºC mp = 45ºC 3 + NaOH density = 1.07 g/mL H2O, H , heat OH CuSO4-(H2O)5 copper(II) sulfate 249.7 g/mol Phenol Side Product Today’s Reaction: Synthesis of p-Chlorotoluene In today’s experiment, we will use copper(I) chloride (CuCl) as our “nucleophile”, to produce the chloride product. The diazonium ion has limited stability in water; it tends to react to give phenols (ArOH) upon standing or unnecessary warming. The principle organic side product will be the phenol resulting from competing reaction with water.