Systems Biology Analyses of Hematopoietic Cells
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(12) United States Patent (10) Patent No.: US 8,603,824 B2 Ramseier Et Al
USOO8603824B2 (12) United States Patent (10) Patent No.: US 8,603,824 B2 Ramseier et al. (45) Date of Patent: Dec. 10, 2013 (54) PROCESS FOR IMPROVED PROTEIN 5,399,684 A 3, 1995 Davie et al. EXPRESSION BY STRAIN ENGINEERING 5,418, 155 A 5/1995 Cormier et al. 5,441,934 A 8/1995 Krapcho et al. (75) Inventors: Thomas M. Ramseier, Poway, CA 5,508,192 A * 4/1996 Georgiou et al. .......... 435/252.3 (US); Hongfan Jin, San Diego, CA 5,527,883 A 6/1996 Thompson et al. (US); Charles H. Squires, Poway, CA 5,558,862 A 9, 1996 Corbinet al. 5,559,015 A 9/1996 Capage et al. (US) 5,571,694 A 11/1996 Makoff et al. (73) Assignee: Pfenex, Inc., San Diego, CA (US) 5,595,898 A 1/1997 Robinson et al. 5,610,044 A 3, 1997 Lam et al. (*) Notice: Subject to any disclaimer, the term of this 5,621,074 A 4/1997 Bjorn et al. patent is extended or adjusted under 35 5,622,846 A 4/1997 Kiener et al. 5,641,671 A 6/1997 Bos et al. U.S.C. 154(b) by 471 days. 5,641,870 A 6/1997 Rinderknecht et al. 5,643,774 A 7/1997 Ligon et al. (21) Appl. No.: 11/189,375 5,662,898 A 9/1997 Ligon et al. (22) Filed: Jul. 26, 2005 5,677,127 A 10/1997 Hogan et al. 5,683,888 A 1 1/1997 Campbell (65) Prior Publication Data 5,686,282 A 11/1997 Lam et al. -

HNRPH1 (HNRNPH1) (NM 005520) Human Tagged ORF Clone Product Data
OriGene Technologies, Inc. 9620 Medical Center Drive, Ste 200 Rockville, MD 20850, US Phone: +1-888-267-4436 [email protected] EU: [email protected] CN: [email protected] Product datasheet for RC201834 HNRPH1 (HNRNPH1) (NM_005520) Human Tagged ORF Clone Product data: Product Type: Expression Plasmids Product Name: HNRPH1 (HNRNPH1) (NM_005520) Human Tagged ORF Clone Tag: Myc-DDK Symbol: HNRNPH1 Synonyms: hnRNPH; HNRPH; HNRPH1 Vector: pCMV6-Entry (PS100001) E. coli Selection: Kanamycin (25 ug/mL) Cell Selection: Neomycin This product is to be used for laboratory only. Not for diagnostic or therapeutic use. View online » ©2021 OriGene Technologies, Inc., 9620 Medical Center Drive, Ste 200, Rockville, MD 20850, US 1 / 6 HNRPH1 (HNRNPH1) (NM_005520) Human Tagged ORF Clone – RC201834 ORF Nucleotide >RC201834 ORF sequence Sequence: Red=Cloning site Blue=ORF Green=Tags(s) TTTTGTAATACGACTCACTATAGGGCGGCCGGGAATTCGTCGACTGGATCCGGTACCGAGGAGATCTGCC GCCGCGATCGCC ATGATGTTGGGCACGGAAGGTGGAGAGGGATTCGTGGTGAAGGTCCGGGGCTTGCCCTGGTCTTGCTCGG CCGATGAAGTGCAGAGGTTTTTTTCTGACTGCAAAATTCAAAATGGGGCTCAAGGTATTCGTTTCATCTA CACCAGAGAAGGCAGACCAAGTGGCGAGGCTTTTGTTGAACTTGAATCAGAAGATGAAGTCAAATTGGCC CTGAAAAAAGACAGAGAAACTATGGGACACAGATATGTTGAAGTATTCAAGTCAAACAACGTTGAAATGG ATTGGGTGTTGAAGCATACTGGTCCAAATAGTCCTGACACGGCCAATGATGGCTTTGTACGGCTTAGAGG ACTTCCCTTTGGATGTAGCAAGGAAGAAATTGTTCAGTTCTTCTCAGGGTTGGAAATCGTGCCAAATGGG ATAACATTGCCGGTGGACTTCCAGGGGAGGAGTACGGGGGAGGCCTTCGTGCAGTTTGCTTCACAGGAAA TAGCTGAAAAGGCTCTAAAGAAACACAAGGAAAGAATAGGGCACAGGTATATTGAAATCTTTAAGAGCAG TAGAGCTGAAGTTAGAACTCATTATGATCCACCACGAAAGCTTATGGCCATGCAGCGGCCAGGTCCTTAT -

Snapshot: the Splicing Regulatory Machinery Mathieu Gabut, Sidharth Chaudhry, and Benjamin J
192 Cell SnapShot: The Splicing Regulatory Machinery Mathieu Gabut, Sidharth Chaudhry, and Benjamin J. Blencowe 133 Banting and Best Department of Medical Research, University of Toronto, Toronto, ON M5S 3E1, Canada Expression in mouse , April4, 2008©2008Elsevier Inc. Low High Name Other Names Protein Domains Binding Sites Target Genes/Mouse Phenotypes/Disease Associations Amy Ceb Hip Hyp OB Eye SC BM Bo Ht SM Epd Kd Liv Lu Pan Pla Pro Sto Spl Thy Thd Te Ut Ov E6.5 E8.5 E10.5 SRp20 Sfrs3, X16 RRM, RS GCUCCUCUUC SRp20, CT/CGRP; −/− early embryonic lethal E3.5 9G8 Sfrs7 RRM, RS, C2HC Znf (GAC)n Tau, GnRH, 9G8 ASF/SF2 Sfrs1 RRM, RS RGAAGAAC HipK3, CaMKIIδ, HIV RNAs; −/− embryonic lethal, cond. KO cardiomyopathy SC35 Sfrs2 RRM, RS UGCUGUU AChE; −/− embryonic lethal, cond. KO deficient T-cell maturation, cardiomyopathy; LS SRp30c Sfrs9 RRM, RS CUGGAUU Glucocorticoid receptor SRp38 Fusip1, Nssr RRM, RS ACAAAGACAA CREB, type II and type XI collagens SRp40 Sfrs5, HRS RRM, RS AGGAGAAGGGA HipK3, PKCβ-II, Fibronectin SRp55 Sfrs6 RRM, RS GGCAGCACCUG cTnT, CD44 DOI 10.1016/j.cell.2008.03.010 SRp75 Sfrs4 RRM, RS GAAGGA FN1, E1A, CD45; overexpression enhances chondrogenic differentiation Tra2α Tra2a RRM, RS GAAARGARR GnRH; overexpression promotes RA-induced neural differentiation SR and SR-Related Proteins Tra2β Sfrs10 RRM, RS (GAA)n HipK3, SMN, Tau SRm160 Srrm1 RS, PWI AUGAAGAGGA CD44 SWAP Sfrs8 RS, SWAP ND SWAP, CD45, Tau; possible asthma susceptibility gene hnRNP A1 Hnrnpa1 RRM, RGG UAGGGA/U HipK3, SMN2, c-H-ras; rheumatoid arthritis, systemic lupus -

AF0033-Phospho-Hnrpd (Ser83) Antibody
Affinity Biosciences website:www.affbiotech.com order:[email protected] Phospho-hnRPD (Ser83) Antibody Cat.#: AF0033 Concn.: 1mg/ml Mol.Wt.: 38kDa Size: 100ul,200ul Source: Rabbit Clonality: Polyclonal Application: WB 1:500-1:2000, IHC 1:50-1:200, IF/ICC 1:100-1:500, ELISA(peptide) 1:20000-1:40000 *The optimal dilutions should be determined by the end user. Reactivity: Human,Mouse,Rat Purification: The antibody is from purified rabbit serum by affinity purification via sequential chromatography on phospho- peptide and non-phospho-peptide affinity columns. Specificity: Phospho-hnRPD (Ser83) Antibody detects endogenous levels of hnRPD only when phosphorylated at Sersine 83. Immunogen: A synthesized peptide derived from human hnRPD around the phosphorylation site of Ser83. Uniprot: Q14103 Description: This gene belongs to the subfamily of ubiquitously expressed heterogeneous nuclear ribonucleoproteins (hnRNPs). The hnRNPs are nucleic acid binding proteins and they complex with heterogeneous nuclear RNA (hnRNA). These proteins are associated with pre-mRNAs in the nucleus and appear to influence pre-mRNA processing and other aspects of mRNA metabolism and transport. While all of the hnRNPs are present in the nucleus, some seem to shuttle between the nucleus and the cytoplasm. The hnRNP proteins have distinct nucleic acid binding properties. The protein encoded by this gene has two repeats of quasi-RRM domains that bind to RNAs. It localizes to both the nucleus and the cytoplasm. This protein is implicated in the regulation of mRNA stability. Alternative splicing of this gene results in four transcript variants. Storage Condition and Rabbit IgG in phosphate buffered saline , pH 7.4, 150mM Buffer: NaCl, 0.02% sodium azide and 50% glycerol.Store at -20 °C.Stable for 12 months from date of receipt. -

Supplementary Table S4. FGA Co-Expressed Gene List in LUAD
Supplementary Table S4. FGA co-expressed gene list in LUAD tumors Symbol R Locus Description FGG 0.919 4q28 fibrinogen gamma chain FGL1 0.635 8p22 fibrinogen-like 1 SLC7A2 0.536 8p22 solute carrier family 7 (cationic amino acid transporter, y+ system), member 2 DUSP4 0.521 8p12-p11 dual specificity phosphatase 4 HAL 0.51 12q22-q24.1histidine ammonia-lyase PDE4D 0.499 5q12 phosphodiesterase 4D, cAMP-specific FURIN 0.497 15q26.1 furin (paired basic amino acid cleaving enzyme) CPS1 0.49 2q35 carbamoyl-phosphate synthase 1, mitochondrial TESC 0.478 12q24.22 tescalcin INHA 0.465 2q35 inhibin, alpha S100P 0.461 4p16 S100 calcium binding protein P VPS37A 0.447 8p22 vacuolar protein sorting 37 homolog A (S. cerevisiae) SLC16A14 0.447 2q36.3 solute carrier family 16, member 14 PPARGC1A 0.443 4p15.1 peroxisome proliferator-activated receptor gamma, coactivator 1 alpha SIK1 0.435 21q22.3 salt-inducible kinase 1 IRS2 0.434 13q34 insulin receptor substrate 2 RND1 0.433 12q12 Rho family GTPase 1 HGD 0.433 3q13.33 homogentisate 1,2-dioxygenase PTP4A1 0.432 6q12 protein tyrosine phosphatase type IVA, member 1 C8orf4 0.428 8p11.2 chromosome 8 open reading frame 4 DDC 0.427 7p12.2 dopa decarboxylase (aromatic L-amino acid decarboxylase) TACC2 0.427 10q26 transforming, acidic coiled-coil containing protein 2 MUC13 0.422 3q21.2 mucin 13, cell surface associated C5 0.412 9q33-q34 complement component 5 NR4A2 0.412 2q22-q23 nuclear receptor subfamily 4, group A, member 2 EYS 0.411 6q12 eyes shut homolog (Drosophila) GPX2 0.406 14q24.1 glutathione peroxidase -

Placenta-Derived Exosomes Continuously Increase in Maternal
Sarker et al. Journal of Translational Medicine 2014, 12:204 http://www.translational-medicine.com/content/12/1/204 RESEARCH Open Access Placenta-derived exosomes continuously increase in maternal circulation over the first trimester of pregnancy Suchismita Sarker1, Katherin Scholz-Romero1, Alejandra Perez2, Sebastian E Illanes1,2,3, Murray D Mitchell1, Gregory E Rice1,2 and Carlos Salomon1,2* Abstract Background: Human placenta releases specific nanovesicles (i.e. exosomes) into the maternal circulation during pregnancy, however, the presence of placenta-derived exosomes in maternal blood during early pregnancy remains to be established. The aim of this study was to characterise gestational age related changes in the concentration of placenta-derived exosomes during the first trimester of pregnancy (i.e. from 6 to 12 weeks) in plasma from women with normal pregnancies. Methods: A time-series experimental design was used to establish pregnancy-associated changes in maternal plasma exosome concentrations during the first trimester. A series of plasma were collected from normal healthy women (10 patients) at 6, 7, 8, 9, 10, 11 and 12 weeks of gestation (n = 70). We measured the stability of these vesicles by quantifying and observing their protein and miRNA contents after the freeze/thawing processes. Exosomes were isolated by differential and buoyant density centrifugation using a sucrose continuous gradient and characterised by their size distribution and morphology using the nanoparticles tracking analysis (NTA; Nanosight™) and electron microscopy (EM), respectively. The total number of exosomes and placenta-derived exosomes were determined by quantifying the immunoreactive exosomal marker, CD63 and a placenta-specific marker (Placental Alkaline Phosphatase PLAP). -

(P -Value<0.05, Fold Change≥1.4), 4 Vs. 0 Gy Irradiation
Table S1: Significant differentially expressed genes (P -Value<0.05, Fold Change≥1.4), 4 vs. 0 Gy irradiation Genbank Fold Change P -Value Gene Symbol Description Accession Q9F8M7_CARHY (Q9F8M7) DTDP-glucose 4,6-dehydratase (Fragment), partial (9%) 6.70 0.017399678 THC2699065 [THC2719287] 5.53 0.003379195 BC013657 BC013657 Homo sapiens cDNA clone IMAGE:4152983, partial cds. [BC013657] 5.10 0.024641735 THC2750781 Ciliary dynein heavy chain 5 (Axonemal beta dynein heavy chain 5) (HL1). 4.07 0.04353262 DNAH5 [Source:Uniprot/SWISSPROT;Acc:Q8TE73] [ENST00000382416] 3.81 0.002855909 NM_145263 SPATA18 Homo sapiens spermatogenesis associated 18 homolog (rat) (SPATA18), mRNA [NM_145263] AA418814 zw01a02.s1 Soares_NhHMPu_S1 Homo sapiens cDNA clone IMAGE:767978 3', 3.69 0.03203913 AA418814 AA418814 mRNA sequence [AA418814] AL356953 leucine-rich repeat-containing G protein-coupled receptor 6 {Homo sapiens} (exp=0; 3.63 0.0277936 THC2705989 wgp=1; cg=0), partial (4%) [THC2752981] AA484677 ne64a07.s1 NCI_CGAP_Alv1 Homo sapiens cDNA clone IMAGE:909012, mRNA 3.63 0.027098073 AA484677 AA484677 sequence [AA484677] oe06h09.s1 NCI_CGAP_Ov2 Homo sapiens cDNA clone IMAGE:1385153, mRNA sequence 3.48 0.04468495 AA837799 AA837799 [AA837799] Homo sapiens hypothetical protein LOC340109, mRNA (cDNA clone IMAGE:5578073), partial 3.27 0.031178378 BC039509 LOC643401 cds. [BC039509] Homo sapiens Fas (TNF receptor superfamily, member 6) (FAS), transcript variant 1, mRNA 3.24 0.022156298 NM_000043 FAS [NM_000043] 3.20 0.021043295 A_32_P125056 BF803942 CM2-CI0135-021100-477-g08 CI0135 Homo sapiens cDNA, mRNA sequence 3.04 0.043389246 BF803942 BF803942 [BF803942] 3.03 0.002430239 NM_015920 RPS27L Homo sapiens ribosomal protein S27-like (RPS27L), mRNA [NM_015920] Homo sapiens tumor necrosis factor receptor superfamily, member 10c, decoy without an 2.98 0.021202829 NM_003841 TNFRSF10C intracellular domain (TNFRSF10C), mRNA [NM_003841] 2.97 0.03243901 AB002384 C6orf32 Homo sapiens mRNA for KIAA0386 gene, partial cds. -

Appendix 2. Significantly Differentially Regulated Genes in Term Compared with Second Trimester Amniotic Fluid Supernatant
Appendix 2. Significantly Differentially Regulated Genes in Term Compared With Second Trimester Amniotic Fluid Supernatant Fold Change in term vs second trimester Amniotic Affymetrix Duplicate Fluid Probe ID probes Symbol Entrez Gene Name 1019.9 217059_at D MUC7 mucin 7, secreted 424.5 211735_x_at D SFTPC surfactant protein C 416.2 206835_at STATH statherin 363.4 214387_x_at D SFTPC surfactant protein C 295.5 205982_x_at D SFTPC surfactant protein C 288.7 1553454_at RPTN repetin solute carrier family 34 (sodium 251.3 204124_at SLC34A2 phosphate), member 2 238.9 206786_at HTN3 histatin 3 161.5 220191_at GKN1 gastrokine 1 152.7 223678_s_at D SFTPA2 surfactant protein A2 130.9 207430_s_at D MSMB microseminoprotein, beta- 99.0 214199_at SFTPD surfactant protein D major histocompatibility complex, class II, 96.5 210982_s_at D HLA-DRA DR alpha 96.5 221133_s_at D CLDN18 claudin 18 94.4 238222_at GKN2 gastrokine 2 93.7 1557961_s_at D LOC100127983 uncharacterized LOC100127983 93.1 229584_at LRRK2 leucine-rich repeat kinase 2 HOXD cluster antisense RNA 1 (non- 88.6 242042_s_at D HOXD-AS1 protein coding) 86.0 205569_at LAMP3 lysosomal-associated membrane protein 3 85.4 232698_at BPIFB2 BPI fold containing family B, member 2 84.4 205979_at SCGB2A1 secretoglobin, family 2A, member 1 84.3 230469_at RTKN2 rhotekin 2 82.2 204130_at HSD11B2 hydroxysteroid (11-beta) dehydrogenase 2 81.9 222242_s_at KLK5 kallikrein-related peptidase 5 77.0 237281_at AKAP14 A kinase (PRKA) anchor protein 14 76.7 1553602_at MUCL1 mucin-like 1 76.3 216359_at D MUC7 mucin 7, -

Pirfenidone Is Renoprotective in Diabetic Kidney Disease
BASIC RESEARCH www.jasn.org Pirfenidone Is Renoprotective in Diabetic Kidney Disease ʈ Satish P. RamachandraRao,*†‡ Yanqing Zhu,‡ Timothy Ravasi,§ Tracy A. McGowan,‡ Irene Toh,‡ Stephen R. Dunn,‡¶ Shinichi Okada,*† Michael A. Shaw,** and Kumar Sharma*†‡ *Center for Renal Translational Medicine, Division of Nephrology-Hypertension, Department of Medicine, and ʈ §Department of Bioengineering, Jacobs School of Engineering, University of California, San Diego, Scripps NeuroAIDS Preclinical Studies Centre, and †Veterans Administration San Diego Healthcare System, La Jolla, California, ‡Center for Novel Therapies in Kidney Disease, Department of Medicine, ¶Cancer Genomics Facility, Kimmel Cancer Center, and **Proteomics and Mass Spectrometry Core Facility, Department of Cancer Biology, Thomas Jefferson University, Philadelphia, Pennsylvania ABSTRACT Although several interventions slow the progression of diabetic nephropathy, current therapies do not halt progression completely. Recent preclinical studies suggested that pirfenidone (PFD) prevents fibrosis in various diseases, but the mechanisms underlying its antifibrotic action are incompletely understood. Here, we evaluated the role of PFD in regulation of the extracellular matrix. In mouse mesangial cells, PFD decreased TGF- promoter activity, reduced TGF- protein secretion, and inhibited TGF-–induced Smad2-phosphor- ylation, 3TP-lux promoter activity, and generation of reactive oxygen species. To explore the therapeutic potential of PFD, we administered PFD to 17-wk-old db/db mice for 4 wk. PFD treatment significantly reduced mesangial matrix expansion and expression of renal matrix genes but did not affect albuminuria. Using liquid chromatography with subsequent electrospray ionization tandem mass spectrometry, we iden- tified 21 proteins unique to PFD-treated diabetic kidneys. Analysis of gene ontology and protein–protein interactions of these proteins suggested that PFD may regulate RNA processing. -

(12) Patent Application Publication (10) Pub. No.: US 2006/0110747 A1 Ramseier Et Al
US 200601 10747A1 (19) United States (12) Patent Application Publication (10) Pub. No.: US 2006/0110747 A1 Ramseier et al. (43) Pub. Date: May 25, 2006 (54) PROCESS FOR IMPROVED PROTEIN (60) Provisional application No. 60/591489, filed on Jul. EXPRESSION BY STRAIN ENGINEERING 26, 2004. (75) Inventors: Thomas M. Ramseier, Poway, CA Publication Classification (US); Hongfan Jin, San Diego, CA (51) Int. Cl. (US); Charles H. Squires, Poway, CA CI2O I/68 (2006.01) (US) GOIN 33/53 (2006.01) CI2N 15/74 (2006.01) Correspondence Address: (52) U.S. Cl. ................................ 435/6: 435/7.1; 435/471 KING & SPALDING LLP 118O PEACHTREE STREET (57) ABSTRACT ATLANTA, GA 30309 (US) This invention is a process for improving the production levels of recombinant proteins or peptides or improving the (73) Assignee: Dow Global Technologies Inc., Midland, level of active recombinant proteins or peptides expressed in MI (US) host cells. The invention is a process of comparing two genetic profiles of a cell that expresses a recombinant (21) Appl. No.: 11/189,375 protein and modifying the cell to change the expression of a gene product that is upregulated in response to the recom (22) Filed: Jul. 26, 2005 binant protein expression. The process can improve protein production or can improve protein quality, for example, by Related U.S. Application Data increasing solubility of a recombinant protein. Patent Application Publication May 25, 2006 Sheet 1 of 15 US 2006/0110747 A1 Figure 1 09 010909070£020\,0 10°0 Patent Application Publication May 25, 2006 Sheet 2 of 15 US 2006/0110747 A1 Figure 2 Ester sers Custer || || || || || HH-I-H 1 H4 s a cisiers TT closers | | | | | | Ya S T RXFO 1961. -
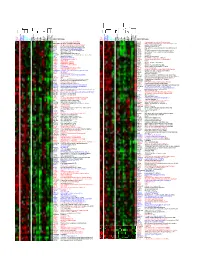
Supporting Figure 8
HUGO ID Name HUGO ID Name Nalm-6 TOM-1 Reh Karpas-422 DoHH-2 SU-DHL-5 Nama lwa DG-75 Ramos Raji BEL EHEB BONNA-12 L-428 DEL BCP-1 BC-3 BCBL-1 JSC-1 PEL -SY HBL-6 DS-1 RPMI-8226 NCI-H929 L-363 SK-MM-2 Nalm-6 TOM-1 Reh Karpas-422 DoHH-2 SU-DHL-5 Nama lwa DG-75 Ramos Raji BEL EHEB BONNA-12 L-428 DEL BCP-1 BC-3 BCBL-1 JSC-1 PEL -SY HBL-6 DS-1 RPMI-8226 NCI-H929 L-363 SK-MM-2 CDC5L Cdc5-related protein (PCDC5RP) RAD51 Replication protein A (E coli RecA homolog) HSU53209 transformer-2 alpha (htra-2 alpha) HMG2 High-mobility group (nonhistone chromosomal) protein 2 CDC10 similar to TR:G560623 G560623 HCDC10 SPS selenium donor protein (selD) NFYB CAAT-box DNA binding protein subunit B CES2 Carboxylestease 2 (liver) GTF2I Bruton's tyrosine kinase-associated protein NOE1 clone 23876 neuronal olfactomedin-related ER localized CCT4 stimulator of TAR RNA binding (SRB) ZNF22 HKR-T1 RY1 RY-1 putative nucleic acid binding protein FKBP5 54 kDa progesterone receptor-associated FKBP54 DVL2 dishevelled 2 (DVL2) FKBP5 54 kDa progesterone receptor-associated FKBP54 RPL13 60S RIBOSOMAL PROTEIN L13 CU L4 A Hs-cul-4A C14ORF3 Hydroxyacyl-Coenzyme A dehydrogenase, beta subuit CU L4 A Hs-cul-4A SM S spermine synthase CLCN6 putative chloride channel SF3A1 splicing factor SF3a120 FALZ fetal Alz-50-reactive clone 1 (FAC1) KNSL4 oriP binding protein (OBP-1) GTF2H3 General transcription factor IIH, polypeptide 3 TUBA1 alpha-tubulin TRIP hT RIP TUBA1 Alpha-tubulin (K00558) GM2A glucose transporter pseudogene TUBA1 Alpha-tubulin (K00558) RBBP6 RBQ-1 TUBA1 TUBULIN ALPHA-4 -

Evolutionary Fate of Retroposed Gene Copies in the Human Genome
Evolutionary fate of retroposed gene copies in the human genome Nicolas Vinckenbosch*, Isabelle Dupanloup*†, and Henrik Kaessmann*‡ *Center for Integrative Genomics, University of Lausanne, Ge´nopode, 1015 Lausanne, Switzerland; and †Computational and Molecular Population Genetics Laboratory, Zoological Institute, University of Bern, 3012 Bern, Switzerland Communicated by Wen-Hsiung Li, University of Chicago, Chicago, IL, December 30, 2005 (received for review December 14, 2005) Given that retroposed copies of genes are presumed to lack the and rodent genomes (7–12). In addition, three recent studies regulatory elements required for their expression, retroposition using EST data (13, 14) and tiling-microarray data from chro- has long been considered a mechanism without functional rele- mosome 22 (15) indicated that retrocopy transcription may be vance. However, through an in silico assay for transcriptional widespread, although these surveys were limited, and potential activity, we identify here >1,000 transcribed retrocopies in the functional implications were not addressed. human genome, of which at least Ϸ120 have evolved into bona To further explore the functional significance of retroposition fide genes. Among these, Ϸ50 retrogenes have evolved functions in the human genome, we systematically screened for signatures in testes, more than half of which were recruited as functional of selection related to retrocopy transcription. Our results autosomal counterparts of X-linked genes during spermatogene- suggest that retrocopy transcription is not rare and that the sis. Generally, retrogenes emerge ‘‘out of the testis,’’ because they pattern of transcription of human retrocopies has been pro- are often initially transcribed in testis and later evolve stronger and foundly shaped by natural selection, acting both for and against sometimes more diverse spatial expression patterns.