Colour Vision 2 Metamers and Theories of Colour Perception
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Chapter 6 COLOR and COLOR VISION
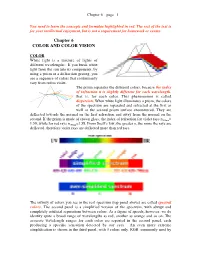
Chapter 6 – page 1 You need to learn the concepts and formulae highlighted in red. The rest of the text is for your intellectual enjoyment, but is not a requirement for homework or exams. Chapter 6 COLOR AND COLOR VISION COLOR White light is a mixture of lights of different wavelengths. If you break white light from the sun into its components, by using a prism or a diffraction grating, you see a sequence of colors that continuously vary from red to violet. The prism separates the different colors, because the index of refraction n is slightly different for each wavelength, that is, for each color. This phenomenon is called dispersion. When white light illuminates a prism, the colors of the spectrum are separated and refracted at the first as well as the second prism surface encountered. They are deflected towards the normal on the first refraction and away from the normal on the second. If the prism is made of crown glass, the index of refraction for violet rays n400nm= 1.59, while for red rays n700nm=1.58. From Snell’s law, the greater n, the more the rays are deflected, therefore violet rays are deflected more than red rays. The infinity of colors you see in the real spectrum (top panel above) are called spectral colors. The second panel is a simplified version of the spectrum, with abrupt and completely artificial separations between colors. As a figure of speech, however, we do identify quite a broad range of wavelengths as red, another as orange and so on. -

Colour Vision Deficiency
Eye (2010) 24, 747–755 & 2010 Macmillan Publishers Limited All rights reserved 0950-222X/10 $32.00 www.nature.com/eye Colour vision MP Simunovic REVIEW deficiency Abstract effective "treatment" of colour vision deficiency: whilst it has been suggested that tinted lenses Colour vision deficiency is one of the could offer a means of enabling those with commonest disorders of vision and can be colour vision deficiency to make spectral divided into congenital and acquired forms. discriminations that would normally elude Congenital colour vision deficiency affects as them, clinical trials of such lenses have been many as 8% of males and 0.5% of femalesFthe largely disappointing. Recent developments in difference in prevalence reflects the fact that molecular genetics have enabled us to not only the commonest forms of congenital colour understand more completely the genetic basis of vision deficiency are inherited in an X-linked colour vision deficiency, they have opened the recessive manner. Until relatively recently, our possibility of gene therapy. The application of understanding of the pathophysiological basis gene therapy to animal models of colour vision of colour vision deficiency largely rested on deficiency has shown dramatic results; behavioural data; however, modern molecular furthermore, it has provided interesting insights genetic techniques have helped to elucidate its into the plasticity of the visual system with mechanisms. respect to extracting information about the The current management of congenital spectral composition of the visual scene. colour vision deficiency lies chiefly in appropriate counselling (including career counselling). Although visual aids may Materials and methods be of benefit to those with colour vision deficiency when performing certain tasks, the This article was prepared by performing a evidence suggests that they do not enable primary search of Pubmed for articles on wearers to obtain normal colour ‘colo(u)r vision deficiency’ and ‘colo(u)r discrimination. -

The Neural Control of Fast Vs. Slow Vergence Eye Movements
European Journal of Neuroscience European Journal of Neuroscience, Vol. 33, pp. 2147–2154, 2011 doi:10.1111/j.1460-9568.2011.07692.x The neural control of fast vs. slow vergence eye movements Kathleen E. Cullen and Marion R. Van Horn Department of Physiology, Aerospace Medical Research Unit, McGill University, Montreal, PQ, Canada Keywords: binocular, burst neuron, conjugate, extraocular, monocular, saccade Abstract When looking between targets located in three-dimensional space, information about relative depth is sent from the visual cortex to the motor control centers in the brainstem, which are responsible for generating appropriate motor commands to move the eyes. Surprisingly, how the neurons in the brainstem use the depth information supplied by the visual cortex to precisely aim each eye on a visual target remains highly controversial. This review will consider the results of recent studies that have focused on determining how individual neurons contribute to realigning gaze when we look between objects located at different depths. In particular, the results of new experiments provide compelling evidence that the majority of saccadic neurons dynamically encode the movement of an individual eye, and show that the time-varying discharge of the saccadic neuron population encodes the drive required to account for vergence facilitation during disconjugate saccades. Notably, these results suggest that an additional input (i.e. from a separate vergence subsystem) is not required to shape the activity of motoneurons during disconjugate saccades. Furthermore, whereas motoneurons drive both fast and slow vergence movements, saccadic neurons discharge only during fast vergence movements, emphasizing the existence of distinct premotor pathways for controlling fast vs. -

1 Human Color Vision
CAMC01 9/30/04 3:13 PM Page 1 1 Human Color Vision Color appearance models aim to extend basic colorimetry to the level of speci- fying the perceived color of stimuli in a wide variety of viewing conditions. To fully appreciate the formulation, implementation, and application of color appearance models, several fundamental topics in color science must first be understood. These are the topics of the first few chapters of this book. Since color appearance represents several of the dimensions of our visual experience, any system designed to predict correlates to these experiences must be based, to some degree, on the form and function of the human visual system. All of the color appearance models described in this book are derived with human visual function in mind. It becomes much simpler to understand the formulations of the various models if the basic anatomy, physiology, and performance of the visual system is understood. Thus, this book begins with a treatment of the human visual system. As necessitated by the limited scope available in a single chapter, this treatment of the visual system is an overview of the topics most important for an appreciation of color appearance modeling. The field of vision science is immense and fascinating. Readers are encouraged to explore the liter- ature and the many useful texts on human vision in order to gain further insight and details. Of particular note are the review paper on the mechan- isms of color vision by Lennie and D’Zmura (1988), the text on human color vision by Kaiser and Boynton (1996), the more general text on the founda- tions of vision by Wandell (1995), the comprehensive treatment by Palmer (1999), and edited collections on color vision by Backhaus et al. -

Color Vision Deficiency
Color Vision Deficiency What is color vision deficiency? Color vision deficiency is called “color blindness” by mistake. Actually, the term describes a number of different problems people have with color vision. Abnormal color vision may vary from not being able to tell certain colors apart to not being able to identify any color. Whom does color vision deficiency affect? An estimated 8% of males and fewer than 1% of females have color vision problems. Most color vision problems run in families and are inherited and present at birth. A child inherits a color vision deficiency by receiving a faulty color vision gene from a parent. Abnormal color vision is found in a recessive gene on the X chromosome. Men are born with just one X and one Y chromosome. However, women have two X chromosomes. Because of this, women can sometimes overcome the faulty gene with their second normal X chromosome. Men, unfortunately, do not have a second X chromosome to help compensate for the faulty color vision gene. Heredity does not cause all color vision problems. One common problem happens from the normal aging of the eye’s lens. The lens is clear at birth, but the aging process causes it to darken and yellow. Older adults may have problems identifying certain dark colors, particularly blues. Certain medications as well as inherited or acquired retinal and optic nerve disease, may also affect normal color vision. Who should be tested for color deficiency? Any child who is having difficulty in school should be checked for possible visual problems including color vision impairment. -

Our Senses & Information Processing
Our senses What we see Light entering our eyes Question: How Tall does a mirror have to be to see your body from head to toe? Prediction A plane mirror is affixed to the wall. What do you think the minimum length of the mirror must be for you to see your entire body from head to toe (just as tall, ¾ as tall, ½ as tall etc…)? At what height does a mirror of this length need to be mounted to allow you to see your entire body from head to toe? Why is this? Procedure Materials: Plane mirror mounted to the wall, meter stick, dry erase marker. Note: While you’re waiting your turn for the mirror, skip to page 5 and start on one of the activities listed on pages 5, 6 or 7. 1. Choose your two most extreme group members, the tallest and the shortest. A. Have the shortest member, known as the object, stand in front of the plane mirror. Another group member will mark on the mirror with a dry erase marker where the object sees the top of their head. Then mark where the object sees their feet. Cover the unused areas of the mirror to verify that they are not needed for the object to see themselves from head to foot. B. Now have the object test to see how distance from the mirror affects the necessary size of mirror. The object should stand very close and very far from the mirror the entire time checking to see how the original head and feet marks align with their head and feet respectively. -

Vergence Eye Movements Redefined: the Neural Control of Fast Versus Slow Vergence
Vergence eye movements redefined: The neural control of fast versus slow vergence Marion R. Van Horn Aerospace Medical Research Unit Department of Physiology McGill University Montreal, Quebec, Canada August 2010 A thesis submitted to the faculty of Graduate Studies and Research in partial fulfilment of the degree of Doctorate in Philosophy Copyright © Marion Van Horn 2010 TABLE OF CONTENTS ABSTRACT…………………………..…………………………..………………... xiii RÉSUMÉ…………………………..…………………………..…………………… xv ACKNOWLEDGEMENTS…………………………..………………………… ..xviii CONTRIBUTIONS OF AUTHORS…………………………..………………….. xx LIST OF ABBREVIATIONS…………………………………..……………….. xxii CHAPTER 1: GENERAL INTRODUCTION & LITERATURE REVIEW 1.1 Overview and conceptual framework………...……………………………… 1 1.2 The extraocular eye muscles and their innervations…………….…………… 4 1.3 Classification of eye movements………………….…………….…………… 8 1.4 The neural control of disconjugate saccades: Hering versus Helmoholtz... 11 1.4.1 Evidence supporting Hering’s Law …………………………….... 11 1.5 The premotor circuitry of conjugate saccades ………………………….... 14 1.5.1 Saccadic burst neurons…………….…………………………….... 15 1.5.2 Omnipause neurons………………..…………………………….... 16 1.5.3 Oculomotor integration ………………………………………….... 17 1.6 The premotor circuitry of saccade-free vergence ……………………….... 19 1.7 Evidence against “Hering’s Law” ……………………………………….... 20 1.8 Research Goals ……………………………….………………………….... 22 CHAPTER 2: DYNAMIC CHARACTERIZATION OF OCULOMOTONEURONS DURING CONJUGATE AND DISCONJUGATE EYE MOVEMENTS 2.1 ABSTRACT ………………………………………………………………. 25 2.2 INTRODUCTION -

Color Vision Mechanisms
11 COLOR VISION MECHANISMS Andrew Stockman Department of Visual Neuroscience UCL Institute of Opthalmology London, United KIngdom David H. Brainard Department of Psychology University of Pennsylvania Philadelphia, Pennsylvania 11.1 GLOSSARY Achromatic mechanism. Hypothetical psychophysical mechanisms, sometimes equated with the luminance mechanism, which respond primarily to changes in intensity. Note that achromatic mech- anisms may have spectrally opponent inputs, in addition to their primary nonopponent inputs. Bezold-Brücke hue shift. The shift in the hue of a stimulus toward either the yellow or blue invariant hues with increasing intensity. Bipolar mechanism. A mechanism, the response of which has two mutually exclusive types of out- put that depend on the balance between its two opposing inputs. Its response is nulled when its two inputs are balanced. Brightness. A perceptual measure of the apparent intensity of lights. Distinct from luminance in the sense that lights that appear equally bright are not necessarily of equal luminance. Cardinal directions. Stimulus directions in a three-dimensional color space that silence two of the three “cardinal mechanisms.” These are the isolating directions for the L+M, L–M, and S–(L+M) mech- anisms. Note that the isolating directions do not necessarily correspond to mechanism directions. Cardinal mechanisms. The second-site bipolar L–M and S–(L+M) chromatic mechanisms and the L+M luminance mechanism. Chromatic discrimination. Discrimination of a chromatic target from another target or back- ground, typically measured at equiluminance. Chromatic mechanism. Hypothetical psychophysical mechanisms that respond to chromatic stimuli, that is, to stimuli modulated at equiluminance. Color appearance. Subjective appearance of the hue, brightness, and saturation of objects or lights. -

Color Vision
Color Vision Lecture 10 Chapter 5, Part A Jonathan Pillow Sensation & Perception (PSY 345 / NEU 325) Princeton University, Fall 2017 1 Exam #1: Thursday 10/19 Format: multiple-choice, fill-in-the-blank, & short answer What to study: - all material from lectures & slides - precept readings (basic gist & findings of each article) (If something appeared only in the book, and not at all in class or precept or slides, you can probably safely ignore it) Review session: tonight @ 7:00 in PNI A30 2 2015 called…. Grey-and-green, or pink-and-white? 3 4 5 • color vision has evolutionary value • lack of color vision ≠ black & white 6 Basic Principles of Color Perception The book says: “Color is not a physical property but a psychophysical property” 7 Basic Principles of Color Perception • Most of the light we see is reflected • Typical light sources: Sun, light bulb, LED screen • We see only part of the electromagnetic spectrum(between 400 and 700 nm). Why?? 8 Basic Principles of Color Perception • Why only 400-700 nm? (Pomerantz, Rice U.) Suggestion: unique ability to penetrate sea water 9 Basic Principles of Color Perception Q: How many numbers would you need to write down to specify the spectral properties of a light source? A: It depends on how you “bin” up the spectrum • One number for each spectral “bin”: 20 17 16 15 13 12 10 example: 13 5 bins energy 0 0 0 0 0 10 Basic Principles of Color Perception Device: hyper-spectral camera - measures the amount of energy (or number of photons) in each small range of wavelengths - can use thousands of bins (or “frequency bands”) instead of just the 13 shown here 20 17 16 15 13 12 10 5 energy 0 0 0 0 0 11 Basic Principles of Color Perception Some terminology for colored light: spectral - referring to the wavelength of light the illuminant - light source power spectrum - this curve. -

Color Vision and Night Vision Chapter Dingcai Cao 10
Retinal Diagnostics Section 2 For additional online content visit http://www.expertconsult.com Color Vision and Night Vision Chapter Dingcai Cao 10 OVERVIEW ROD AND CONE FUNCTIONS Day vision and night vision are two separate modes of visual Differences in the anatomy and physiology (see Chapters 4, perception and the visual system shifts from one mode to the Autofluorescence imaging, and 9, Diagnostic ophthalmic ultra- other based on ambient light levels. Each mode is primarily sound) of the rod and cone systems underlie different visual mediated by one of two photoreceptor classes in the retina, i.e., functions and modes of visual perception. The rod photorecep- cones and rods. In day vision, visual perception is primarily tors are responsible for our exquisite sensitivity to light, operat- cone-mediated and perceptions are chromatic. In other words, ing over a 108 (100 millionfold) range of illumination from near color vision is present in the light levels of daytime. In night total darkness to daylight. Cones operate over a 1011 range of vision, visual perception is rod-mediated and perceptions are illumination, from moonlit night light levels to light levels that principally achromatic. Under dim illuminations, there is no are so high they bleach virtually all photopigments in the cones. obvious color vision and visual perceptions are graded varia- Together the rods and cones function over a 1014 range of illu- tions of light and dark. Historically, color vision has been studied mination. Depending on the relative activity of rods and cones, as the salient feature of day vision and there has been emphasis a light level can be characterized as photopic (cones alone on analysis of cone activities in color vision. -

Color Appearance Models Second Edition
Color Appearance Models Second Edition Mark D. Fairchild Munsell Color Science Laboratory Rochester Institute of Technology, USA Color Appearance Models Wiley–IS&T Series in Imaging Science and Technology Series Editor: Michael A. Kriss Formerly of the Eastman Kodak Research Laboratories and the University of Rochester The Reproduction of Colour (6th Edition) R. W. G. Hunt Color Appearance Models (2nd Edition) Mark D. Fairchild Published in Association with the Society for Imaging Science and Technology Color Appearance Models Second Edition Mark D. Fairchild Munsell Color Science Laboratory Rochester Institute of Technology, USA Copyright © 2005 John Wiley & Sons Ltd, The Atrium, Southern Gate, Chichester, West Sussex PO19 8SQ, England Telephone (+44) 1243 779777 This book was previously publisher by Pearson Education, Inc Email (for orders and customer service enquiries): [email protected] Visit our Home Page on www.wileyeurope.com or www.wiley.com All Rights Reserved. No part of this publication may be reproduced, stored in a retrieval system or transmitted in any form or by any means, electronic, mechanical, photocopying, recording, scanning or otherwise, except under the terms of the Copyright, Designs and Patents Act 1988 or under the terms of a licence issued by the Copyright Licensing Agency Ltd, 90 Tottenham Court Road, London W1T 4LP, UK, without the permission in writing of the Publisher. Requests to the Publisher should be addressed to the Permissions Department, John Wiley & Sons Ltd, The Atrium, Southern Gate, Chichester, West Sussex PO19 8SQ, England, or emailed to [email protected], or faxed to (+44) 1243 770571. This publication is designed to offer Authors the opportunity to publish accurate and authoritative information in regard to the subject matter covered. -

Points of View in the Modern History of Psychology
Points of View in the Modern History of Psychology Edited by Claude E. Buxton Department of Psychology Yale University New Haven, Connecticut 1985 ACADEMIC PRESS, INC. (Harcourt Brace Jovanovich, Publishers) Orlando San Diego New York London Toronto Montreal Sydney Tokyo Passages from the following are reprinted by permission of the publishers: Newell, Α., Duncker on Thinking, in S. Koch & D. Leary (Eds.), A Century of Psychology as Science. Copyright 1985 by McGraw-Hill. Neisser, U., Cognitive Psychology. © 1967 by Prentice-Hall. COPYRIGHT © 1985 BY ACADEMIC PRESS, INC. ALL RIGHTS RESERVED. NO PART OF THIS PUBLICATION MAY BE REPRODUCED OR TRANSMITTED IN ANY FORM OR BY ANY MEANS, ELECTRONIC OR MECHANICAL, INCLUDING PHOTOCOPY, RECORDING, OR ANY INFORMATION STORAGE AND RETRIEVAL SYSTEM, WITHOUT PERMISSION IN WRITING FROM THE PUBLISHER. ACADEMIC PRESS, INC. Orlando, Florida 32887 United Kingdom Edition published by ACADEMIC PRESS INC. (LONDON) LTD. 24-28 Oval Road, London NW1 7DX LIBRARY OF CONGRESS CATALOGING IN PUBLICATION DATA Main entry under title: Points of view in the modern history of psychology. Includes indexes. 1. Psychology— History. I. Buxton, Claude E. BF81.P57 1985 150\9 85-4010 ISBN 0-12-148510-2 (alk. paper) PRINTED IN THE UNITED STATES OF AMERICA 85 86 87 88 9 8 7 6 5 4 3 2 1 Contributors Numbers in parentheses indicate the pages on which the authors' contributions begin. Mitchell G. Ash (295), Department of History, University of Iowa, Iowa City, Iowa 52242 William Bevan (259), John D. and Catherine T. MacArthur Foundation, Chicago, Illinois 60603 Arthur L. Blumenthal (19, 51), Department of Psychology, University of Massachusetts at Boston, Boston, Massachusetts 02125 Claude E.