Vegetation Pattern and Ecology of Siliceous Boulder Snow Beds on Svalbard
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bulletin 2015 Engaged in Arctic Research and in All Areas of the Arctic Region
[IASC] · INTERNATIONAL ARCTIC SCIENCE COMMITTEE The International Arctic Science Committee (IASC) is a non-governmental, international scientific organization. IASC’s mission is to encourage and facilitate cooperation in all aspects of Arctic research, in all countries IASC engaged in Arctic research and in all areas of the Arctic region. Overall, IASC promotes and supports leading-edge multi-disciplinary research in Bulletin 2015 Bulletin order to foster a greater scientific understanding of the Arctic region and its role in the Earth system. TO ACHIEVE THIS MISSION IASC: • Initiates, coordinates and promotes scientific activities at a circumarctic or international level; • Provides mechanisms and instruments to support science development; • Provides objective and independent scientific advice on issues of science in the Arctic and communicates scientific information to the public; • Seeks to ensure that scientific data and information from the Arctic are safeguarded, freely exchangeable and accessible; • Promotes international access to all geographic areas and the sharing of knowledge, logistics and other resources; • Provides for the freedom and ethical conduct of science; • Promotes and involves the next generation of scientists working in the Arctic; and • Promotes bipolar cooperation through interaction with relevant science IASC 2015 organizations. BULLETIN IASC Bulletin 2015 Telegrafenberg A43 - 14473 Potsdam, Germany ISSN: 1654-7594, ISBN: 978-3-9813637-8-4 Phone: +49 331 2882214 | E-mail: [email protected] | www.iasc.info INTERNATIONAL ARCTIC SCIENCE COMMITTEE Telegrafenberg A43, DE - 14473 Potsdam, Germany www.iasc.info INTERNATIONAL ARCTIC SCIENCE COMMITTEE [IASC] · STRUCTURE Representatives of national scientific organizations from all 22 member countries form the IASC Council. The President of IASC is elected by Council, which also elects 4 Vice-Presidents to serve on the Executive Committee. -

Checklist of Lichenicolous Fungi and Lichenicolous Lichens of Svalbard, Including New Species, New Records and Revisions
Herzogia 26 (2), 2013: 323 –359 323 Checklist of lichenicolous fungi and lichenicolous lichens of Svalbard, including new species, new records and revisions Mikhail P. Zhurbenko* & Wolfgang von Brackel Abstract: Zhurbenko, M. P. & Brackel, W. v. 2013. Checklist of lichenicolous fungi and lichenicolous lichens of Svalbard, including new species, new records and revisions. – Herzogia 26: 323 –359. Hainesia bryonorae Zhurb. (on Bryonora castanea), Lichenochora caloplacae Zhurb. (on Caloplaca species), Sphaerellothecium epilecanora Zhurb. (on Lecanora epibryon), and Trimmatostroma cetrariae Brackel (on Cetraria is- landica) are described as new to science. Forty four species of lichenicolous fungi (Arthonia apotheciorum, A. aspicili- ae, A. epiphyscia, A. molendoi, A. pannariae, A. peltigerina, Cercidospora ochrolechiae, C. trypetheliza, C. verrucosar- ia, Dacampia engeliana, Dactylospora aeruginosa, D. frigida, Endococcus fusiger, E. sendtneri, Epibryon conductrix, Epilichen glauconigellus, Lichenochora coppinsii, L. weillii, Lichenopeltella peltigericola, L. santessonii, Lichenostigma chlaroterae, L. maureri, Llimoniella vinosa, Merismatium decolorans, M. heterophractum, Muellerella atricola, M. erratica, Pronectria erythrinella, Protothelenella croceae, Skyttella mulleri, Sphaerellothecium parmeliae, Sphaeropezia santessonii, S. thamnoliae, Stigmidium cladoniicola, S. collematis, S. frigidum, S. leucophlebiae, S. mycobilimbiae, S. pseudopeltideae, Taeniolella pertusariicola, Tremella cetrariicola, Xenonectriella lutescens, X. ornamentata, -

NEW DATA ABOUT MOSSES on the SVALBARD GLACIERS Olga
NEW DATA ABOUT MOSSES ON THE SVALBARD GLACIERS Olga Belkina Polar-Alpine Botanical Garden-Institute, Kola Science Center of the Russian Academy of Sciences, Apatity, Murmansk Province, Russia; e-mail: [email protected] Rapid melting and retreat of glaciers in the Arctic is a cause of sustainable long‐ In Svalbard moss populations were found on 9 glaciers. In 2012, during re‐examination of the populations a few term existence of ablation zone on them. Sometimes these areas are the habitats In 2007 B.R.Mavlyudov collected one specimen on individuals of Bryum cryophilum Mårtensson and some plants of some mosses partly due to good availability of water and cryoconite Bertilbreen (Paludella squarrosa (Hedw.) Brid.) and of Sanionia uncinata were found among H. polare shoots in substratum. 14 species were found in this unusual habitat on Alaska and Iceland: some specimens on Austre Grønfjordbreen (Ceratodon some large cushions. Therefore the next stage of cushion Andreaea rupestris Hedw., Ceratodon purpureus (Hedw.) Brid., Ditrichum purpureus (Hedw.) Brid., Warnstorfia sarmentosa succession had begun –emergence of a di‐ and multi‐species flexicaule (Schwaegr.) Hampe, Pohlia nutans (Hedw.) Lindb., Polytrichum (Wahlenb.) Hedenäs, Sanionia uncinata (Hedw.) community. A similar process was observed earlier on the juniperinum Hedw. (Benninghoff, 1955), Racomitrium fasciculare (Hedw.) Brid. Loeske, Hygrohypnella polare (Lindb.) Ignatov & bone of a mammal that was lying on the same glacier. (=Codriophorus fascicularis (Hedw.) Bendarek‐Ochyra et Ochyra) (Shacklette, Ignatova.). Ceratodon purpureus settled in center of almost spherical 1966), Drepanocladus berggrenii (C.Jens.) Broth. (Heusser, 1972), Racomitrium In 2009 populations of two latter species were studied cushion of Sanionia uncinata on the both butt‐ends of the crispulum var. -

Total of 10 Pages Only May Be Xeroxed
CENTRE FOR NEWFOUNDLAND STUDIES TOTAL OF 10 PAGES ONLY MAY BE XEROXED (Without Author's Permission) ,, l • ...J ..... The Disjunct Bryophyte Element of the Gulf of St. Lawrence Region: Glacial and Postglacial Dispersal and Migrational Histories By @Rene J. Belland B.Sc., M.Sc. A thesis submitted to the School of Graduate Studies in partial fulfilment of the requirements for the degree of Doctor of Philosophy Department of Biology Memorial University of Newfoundland December, 1Q84 St. John's Newfoundland Abstract The Gulf St. Lawrence region has a bryophyte flora of 698 species. Of these 267 (38%) are disjunct to this region from western North America, eastern Asia, or Europe. The Gulf of St. Lawrence and eastern North American distributions of the disjuncts were analysed and their possible migrational and dispersal histories during and after the Last Glaciation (Wisconsin) examined. Based on eastern North American distribution patterns, the disjuncts fell into 22 sub elements supporting five migrational/ dispersal histories or combinations of these : (1) migration from the south, (2) migration from the north, (3) migration from the west, (4) survival in refugia, and (5) introduction by man. The largest groups of disjuncts had eastern North American distributions supporting either survival of bryophytes in Wisconsin ice-free areas of the Gulf of St. Lawrence or postglacial migration to the Gulf from the south. About 26% of the disjuncts have complex histories and their distributions support two histories. These may have migrated to the Gulf from the west and/or north, or from the west and/or survived glaciation in Gulf ice-free areas. -

Census of Breeding Seabirds on the Northwest Coast of Svalbard 1973 and 1978
Census of breeding seabirds on the northwest coast of Svalbard 1973 and 1978 CHRISTIAN KEMPF AND BENO~TSI~ER Kempf, C. & Sittler, B. 1988: Census of breeding seabirds on the northwest coast of Svalbard 1973 and 1978. Polar Research 6, 195-203. The western coast of Svalbard is one of the world's most important seabird regions (Belopol'skij 1961; L0venskiold 1964; Norderhaug et al. 1977), due to the favourable water temperatures, light regime and amounts of mineral salts (Norderhaug et al. 1977). Seabirds have been censused several times in this area (Kristoffersen 1962; Larsen 1965; Dhondt et al. 1969; Voisin 1970; Norderhaug 1974; Sendstad 1978; Alendal et al. 1982). Except for Larsen (1965), there is no comprehensive and quantitative survey of any part of northwestern Svalbard. Further east, Jepsen & Mobrek (1983) surveyed the area between GrPhuken and Nordaustlandet. Recent concern about the potentially detrimental effects of planned oil exploration and increased human activities in the high Arctic has emphasized the need for more information on the ecology in these regions. This paper provides more comprehensive data on seabird populations in northwestern Svalbard, between southern Prins Karls Forland and Verlegenhuken. The results supplement the studies carried out in 1978 and 1979 by Jepsen & Mobzk (1983) between Verlegenhuken and Kong Karls Land. Chrbtian Kempf and Benoit Sittler, Groupe de Recherches en Ecologie Arctique, 68660 Rombach le Franc, France; December 1986 (reuised September 1988). General area description In 1978, a party of 9 people with base camp at Sorgattet (between Magdalenefjorden and The northwest coast of Svalbard is deeply Smeerenburgfjorden) conducted surveys in indented, with rocky peninsulas rising to 300- Kongsfjorden and Albert I Land (9-13 July), 800 m a.s.1. -

The Lichen-Genus Acarospora in Greenland and Spitsbergen
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by NORA - Norwegian Open Research Archives DET KONGELIGE DEPARTEMENT FOR HANDEL, SJØFART, INDUSTRI, HÅNDVERK OG FISKE RI NORGESSVALBAR�OGISHAV�UNDERSØKELSER LEDER: ADOLF HOEL MEDDELELSE Nr. 27 A. H. MAGNUSSON THE LICHEN-GENUS ACAROSPORA IN GREENLAND AND SPITSBERGEN Reprinted from Nyt .Magazin for Naturvidenskaberne B. LXXV, 1935 OSLO I KOMMISJON HOS JACOB DYBWAD 1935 Reprinted from Nyt Magazin for Naturvidenskaberne. Bind 75, Oslo 19il5. The Lichen-Genus Acarospora in Greenland and Spitsbergen. By A. H. MAGNUSSON ( With 7 figures in the text) uring his expedition to North East Greenland in 1929 Prof. D B. Lynge, Oslo, collected a great number of Acarospora species which were sent to me for revision or determination. To this collection was added a considerable n.umber of specimens collected by Th. M. Fries (Th. Fr.) in West Greenland in 1871 and some from Spitsbergen in 1868. Other collections were made in Greenland by P. F. Scholander 1930, and in Spitsbergen by Lynge 1926 and P. F. Scholander 1931. The supposition that the Acarospora-species of Greenland should resemble those of Northern Scandinavia has appeared to be wrong as there were no less than 8 species that could not be identified with those already known. And there seems to be several new species among the material which was too scanty to allow of a certain determination. The specimens collected hy Th. M. Fries belong to Natur historiska Riksmuseet, Stockholm, and those collected by Lynge and Scholander to Botanisk Museum, Oslo. The specimens are arranged here in the order adopted by me in my Monograph of Acarospora 1929. -
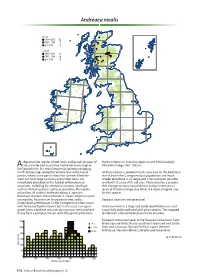
Andreaea Nivalis
Andreaea nivalis Britain 1990–2013 12 1950–1989 7 pre-1950 4 Ireland 1990–2013 0 1950–1989 0 pre-1950 0 characteristic species of wet rocks and gravels in areas of Nardia compressa, Scapania uliginosa and Pohlia ludwigii. Alate snow-lie and associated meltwater burns high in Altitudinal range: 880–1340 m. the Scottish hills. It is most frequently found on dripping, north-facing crags along the ‘cornice line’ at the top of Andreaea nivalis is abundant in its core area on the Ben Nevis corries, where snow persists into the summer. Here the massif and in the Cairngorms but populations are much moss can form large cushions and on Ben Nevis it is smaller elsewhere in its range and it has not been possible remarkably abundant in this habitat with numerous to refind it at some of its old sites. There must be a concern associates, including Deschampsia cespitosa, Saxifraga that changes in snow accumulation and persistence as a stellaris, Anthelia julacea, Lophozia opacifolia, Marsupella result of climate change may affect the more marginal sites sphacelata, M. stableri, Andreaea alpina, A. rupestris, for this species. Ditrichum zonatum, Kiaeria falcata, K. starkei, Polytrichastrum sexangulare, Racomitrium lanuginosum and, rarely, Dioicous; capsules are occasional. Oedipodium griffithianum. In the Cairngorms it often occurs with Andreaea frigida in burns but it also occurs on open Andreaea nivalis is a large and easily identifiable moss and gravel areas which are very wet during snow melt and here is not likely to be confused with other species. The mapped it may form a complex mosaic with Marsupella sphacelata, distribution is therefore believed to be accurate. -

THE ANDEAN SPECIES of PILEA by Ellsworth P. Killip INTRODUCTION
THE ANDEAN SPECIES OF PILEA By Ellsworth P. Killip INTRODUCTION Pilea is by far the largest genus of Urticaceae. Weddell in his final monograph 1 of the family recognized 159 species as valid, about 50 of which were said to occur in the Andes of South America. That these species had a very limited range of distribution was indicated by the fact that more than 30 were known from only a single collection and that only 9 of the others were in any sense widely distributed in the Andean countries. Two, P. microphyUa and P. pubescens, occurred throughout the American Tropics, and P. kyalina and P. dendrophUa extended eastward in South America. In the 50 years following the publication of this monograph only 10 species were described from the Andean region. Because of the large number of specimens of Pilea collected by expeditions to the Andes sponsored by the Smithsonian Institution, the New York Botanical Garden, the Gray Herbarium and the Arnold Arboretum of Harvard University, the Field Museum of Natural History, and the Academy of Natural Sciences, Philadelphia, which were submitted to me for identification but which could not be assigned to any known species, I began the preparation of a mono- graph of all the Andean species. This was completed in 1935 but unfortunately could not be printed in its entirety at that time. Instead, a greatly abridged paper was published,2 which contained a key to all the Andean species, descriptions of 30 new ones, and a few changes of name and of rank. In this there was no opportunity to present descriptions of the earlier species or to discuss their synonymy. -

Chapter 12 Productivity
Glime, J. M. 2017. Productivity. Chapt. 12. In: Glime, J. M. Bryophyte Ecology. Volume 1. Physiological Ecology. Ebook 12-1-1 sponsored by Michigan Technological University and the International Association of Bryologists. Last updated 18 July 2020 and available at <http://digitalcommons.mtu.edu/bryophyte-ecology/>. CHAPTER 12 PRODUCTIVITY TABLE OF CONTENTS Productivity .......................................................................................................................................................... 12-2 Ecological Factors ................................................................................................................................................ 12-2 Ability to Invade ........................................................................................................................................... 12-2 Niche Differences ......................................................................................................................................... 12-3 Growth ................................................................................................................................................................. 12-3 Growth Measurements .................................................................................................................................. 12-4 Annual Length Increase ................................................................................................................................ 12-8 Uncoupling ................................................................................................................................................... -

Shipwreck at Cape Flora: the Expeditions of Benjamin Leigh Smith, England’S Forgotten Arctic Explorer
University of Calgary PRISM: University of Calgary's Digital Repository University of Calgary Press University of Calgary Press Open Access Books 2013 Shipwreck at Cape Flora: The Expeditions of Benjamin Leigh Smith, England’s Forgotten Arctic Explorer Capelotti, P. J. University of Calgary Press Capelotti, P.J. "Shipwreck at Cape Flora: The Expeditions of Benjamin Leigh Smith, England’s Forgotten Arctic Explorer". Northern Lights Series No. 16. University of Calgary Press, Calgary, Alberta, 2013. http://hdl.handle.net/1880/49458 book http://creativecommons.org/licenses/by-nc-nd/3.0/ Attribution Non-Commercial No Derivatives 3.0 Unported Downloaded from PRISM: https://prism.ucalgary.ca University of Calgary Press www.uofcpress.com SHIPWRECK AT CAPE FLORA: THE EXPEDITIONS OF BENJAMIN LEIGH SMITH, ENGLAND’S FORGOTTEN ARCTIC EXPLORER P.J. Capelotti ISBN 978-1-55238-712-2 THIS BOOK IS AN OPEN ACCESS E-BOOK. It is an electronic version of a book that can be purchased in physical form through any bookseller or on-line retailer, or from our distributors. Please support this open access publication by requesting that your university purchase a print copy of this book, or by purchasing a copy yourself. If you have any questions, please contact us at [email protected] Cover Art: The artwork on the cover of this book is not open access and falls under traditional copyright provisions; it cannot be reproduced in any way without written permission of the artists and their agents. The cover can be displayed as a complete cover image for the purposes of publicizing this work, but the artwork cannot be extracted from the context of the cover of this specific work without breaching the artist’s copyright. -

Prioriterte Kulturminner Og Kulturmiljøer Pa Svalbard
KATALOG PRIORITERTE KULTURMINNER OG KULTURMILJØER PA SVALBARD Versjon 1.1 (2013) Irene Skauen Sandodden Sysselmannen på Svalbard Katalog prioriterte kulturminner og kulturmiljøer på Svalbard, versjon 1.1 Side 1 Telefon 79 02 43 00 Internett Adresse Telefaks 79 02 11 66 www.sysselmannen.no Sysselmannen på Svalbard, E-post [email protected] Pb. 633, 9171 Longyearbyen ISBN: Tilgjengelighet Internett: www.sysselmannen.no Opplag: Trykkes ikke, kun digitalt Utgiver Årstall: 2013 Sysselmannen på Svalbard, miljøvernavdelingen Sider: 220 Forfattere Irene Skauen Sandodden. Tekt er hentet fra ulike kilder. Per Kyrre Reymert, Tora Hultgreen, Marit Anne Hauan og Thor Bjørn Arlov har skrevet artikler om de ulike fasene i Svalbard historie (kapittel 2). Deltakende institusjoner Sysselmannen på Svalbard Tittel Title Katalog prioriterte kulturminner og kulturmiljøer på Svalbard. Versjon Catalogue of the cultural heritage sites with high priority in Svalbard. 1.1 (2013). Version 1.1 (2013). Referanse Katalog prioriterte kulturminner og kulturmiljøer på Svalbard. Versjon 1.1 (2013). Tilgjengelig på Internett: www.sysselmannen.no. Sammendrag Katalogen gir et kort innblikk i historien til Svalbard og representative kulturminner. Videre beskrives de om lag 100 prioriterte kulturminnene og kulturmiljøene som er valgt ut i Kulturminneplan for Svalbard 2013 – 2023. Katalogen er utarbeidet som et vedlegg til kulturminneplanen, men kan revideres ved behov. Emneord norsk Keywords English - Kulturminner og kulturmiljø - Cultural heritage (monuments and cultural -

Total of 10 Pages Only May Be Xeroxed
CENTRE FOR NEWFOUNDLAND STUDIES TOTAL OF 10 PAGES ONLY MAY BE XEROXED (Without Author's Permission) I THE BRYOPHYTES OF CATARACTS PROVINCIAL PARK, NEWFOUNDLAND A Thesis Pres en ted to The Department of Biology Hemorial University of Newfoundland In Partial Fulfillment of the Requirements for the Degree Master of Science by David P. Weber © January, 19 76 ii ABSTRACT The bryogeographical knowledge of insular Newfoundland has been significantly augmented through the inten~ investigation of the bryo flora of Cataracts Provincial Park, Avalon Peninsula, and its vicinity. The area investigated included forested areas, bog, heath, and a deep, angular, and narrow river gorge and waterfalls which provide numerous microhabitats particularly suitable for growth of bryophytes. Each bryophyte habitat is fully described, and an annotated list of species gives data on occurrence,frequency,and fertility. Within an area of approximately 13 hectares, 121 species of mosses and 63 species of liverworts were collected, representing one third of the total known bryophyte flora of the island of Newfoundland. The mosses Bryum creberrimum and Ditrichum lineare and the liverwort Cephaloziella hampeana ane reported for the first time from insular Newfoundland. The ranges of a number of Newfoundland bryophytes, including several species previously thought to be more or less restricted to the pre dominantly calcareous regions of the west coast such as Hygrohypnum luridum or Lophozia gillmani, have been extended to include the Avalon Peninsula. A small southern element, represented by Sematophyllum marylandicum, and a northern element, represented by Lophozia bantriensis, are present within the Park but most of the moss and liverwort species there are typical circumboreal species.