An Assessment of Nitrogen and Phosphorus Availability in Constructed Wetlands in the Cumberland Marsh Region, Canada
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Siege of Fort Beauséjour by Chris M. Hand Notes
1 The Siege of Fort Beauséjour by Chris M. Hand Notes Early Conflict in Nova Scotia 1604-1749. By the end of the 1600’s the area was decidedly French. 1713 Treaty of Utrecht After nearly 25 years of continuous war, France ceded Acadia to Britain. French and English disagreed over what actually made up Acadia. The British claimed all of Acadia, the current province of New Brunswick and parts of the current state of Maine. The French conceded Nova Scotia proper but refused to concede what is now New Brunswick and northern Maine, as well as modern Prince Edward Island and Cape Breton. They also chose to limit British ownership along the Chignecto Isthmus and also harboured ambitions to win back the peninsula and most of the Acadian settlers who, after 1713, became subjects of the British Crown. The defacto frontier lay along the Chignecto Isthmus which separates the Bay of Fundy from the Northumberland Strait on the north. Without the Isthmus and the river system to the west, France’s greatest colony along the St. Lawrence River would be completely cut off from November to April. Chignecto was the halfway house between Quebec and Louisbourg. 1721 Paul Mascarene, British governor of Nova Scotia, suggested that a small fort could be built on the neck with a garrison of 150 men. a) one atthe ridge of land at the Acadian town of Beaubassin (now Fort Lawrence) or b) one more west on the more prominent Beauséjour ridge. This never happened because British were busy fighting Mi’kmaq who were incited and abetted by the French. -

The Boundaries of Nationality in Mid-18Th Century Nova Scotia*
GEOFFREY PLANK The Two Majors Cope: The Boundaries of Nationality in Mid-18th Century Nova Scotia* THE 1750S BEGAN OMINOUSLY IN Nova Scotia. In the spring of 1750 a company of French soldiers constructed a fort in a disputed border region on the northern side of the isthmus of Chignecto. The British built a semi-permanent camp only a few hundred yards away. The two armies faced each other nervously, close enough to smell each other's food. In 1754 a similar situation near the Ohio River led to an imperial war. But the empires were not yet ready for war in 1750, and the stand-off at Chignecto lasted five years. i In the early months of the crisis an incident occurred which illustrates many of *' the problems I want to discuss in this essay. On an autumn day in 1750, someone (the identity of this person remains in dispute) approached the British fort waving a white flag. The person wore a powdered wig and the uniform of a French officer. He carried a sword in a sheath by his side. Captain Edward Howe, the commander of the British garrison, responded to the white flag as an invitation to negotiations and went out to greet the man. Then someone, either the man with the flag or a person behind him, shot and killed Captain Howe. According to three near-contemporary accounts of these events, the man in the officer's uniform was not a Frenchman but a Micmac warrior in disguise. He put on the powdered wig and uniform in order to lure Howe out of his fort. -

The Quest for Lobster Stock Boundaries in the Canadian
NOT TO BE CITED WITHOUT PRIOR REFERENCE TO THE AUTHOR(S) Northwest Atlantic Fisheries Organization Serial No. N615 NAFO SCR Doc. 82/IX/107 FOURTH ANNUAL MEETING - SEPTEMBER 1982 The Quest for Lobster Stoclg Boundaries in the Canadian Maritimes by A. Camp ell and R. K. Mohnl Invertebrates and Marine Plants Division Fisheries Research Branch, Department of Fisheries and Oceans Biological Station, St. Andrews, N. B., Canada EOG 2X0 P. O. Box 550, H lifax, N. S., Canada B3J 2S7 Abstract Historical and recent datO on lobsters (Homarus americanus) from the Canadian maritimes were examin4d for stock differences with the object of defining lobster population bot ndaries. Using pattern recognition techniques, historical lobster landings (1892-1981) from 32 areas resulted in grouping the landing trends into 7-8 areas. Examination of morphometrics data, landing trends, population parameters (growth and size at aturity), movement of tagged lobsters, and general surface currents that might indicate larval drift, suggested the following general lobster stock areas: Western maritimes which included the Bay of Fundy, inshore and possibly offshore) southwestner Nova Scotia (to Shelburne Co.), The Eastern Coast of Nova Scotia (Queens to Cape Breton Counties) which seems to be a transition zone f,r lobsters between the Gulf of Maine and the Gulf of St. Lawrence, and 3. Southern Gulf of St. Lawrence STOCK DISCRI INATION SYMPOSIUM Key words: Homarus americanus; lobsters; stocks; population dynamics; morphonietrics; landing statistics; electrophoresis; recruitment; tagging; movement. General Introduction Historically, there have been various debates amongst biologists, fishery managers, and fishermen as to the geographic discreteness of lobster (Homarus americanus) populations on the Atlantic Coast of North America (eg. -

Methodism Among Nova Scotia's Yankee Planters
Methodism Among Nova Scotia's Yankee Planters Allen B. Robertson Queen's University During the 1770s two revivalist evangelical sects gained a following in Nova Scotia; one, Newlight Congregationalism — with both Predestinar- ian and Free Will variants — grew out of the religious and social heritage of the colony's dominant populace, the New England Planters. The other sect, Wesleyan Methodism, took root among transplanted Yorkshiremen who moved between 1772 and 1776 to the Isthmus of Chignecto region where it was initially propagated among the faithful in local prayer groups. Ordained and lay preachers of both movements promoted a series of revivals in the province which drew an increasing number of followers into the evangelical fold.1 The first of these revivals was the Newlight- dominated Great Awakening of 1776-84. In general, Newlightism's greatest appeal was in the Planter townships even though it mutated by 1800 into a Baptist polity. Methodism, which had a fluctuating number of adherents among the visiting military forces at Halifax, had its stronghold in areas settled by British-born colonists, and increased in numbers with the successive waves of Loyalists coming to the province after 1783.2 Methodism was not confined, however, to segregated geographical areas of Nova Scotia. By the early nineteenth century, there were significant Methodist congregations composed primarily of Planters located throughout the Annapolis Valley and along the province's South Shore. Interesting questions are posed for historians when we consider why New Englanders and their descendants were attracted to what appeared to be essentially a foreign hierarchical religious-cultural movement which had broken from 1 Gordon Stewart and George Rawlyk, A People Highly Favoured of God: The Nova Scotia Yankees and the American Revolution (Toronto, 1972); J.M. -

Birds of the Nova Scotia— New Brunswick Border Region by George F
Birds of the Nova Scotia— New Brunswick border region by George F. Boyer Occasional Paper Number 8 Second edition Canadian Wildlife Service Environment Canada Environnement Canada Wildlife Service Service de la Faune Birds of the Nova Scotia - New Brunswick border region by George F. Boyer With addendum by A. J. Erskine and A. D. Smith Canadian Wildlife Service Occasional Paper Number 8 Second edition Issued under the authority of the Honourable Jack Davis, PC, MP Minister of the Environment John S. Tener, Director Canadian Wildlife Service 5 Information Canada, Ottawa, 1972 Catalogue No. CW69-1/8 First edition 1966 Design: Gottschalk-)-Ash Ltd. 4 George Boyer banding a barn swallow in June 1952. The author George Boyer was born in Woodstock, New Brunswick, on August 24, 1916. He graduated in Forestry from the University of New Brunswick in 1938 and served with the Canadian Army from 1939 to 1945. He joined the Canadian Wildlife Service in 1947, and worked out of the Sackville office until 1956. During that time he obtained an M.S. in zoology from the University of Illinois. He car ried on private research from April 1956 until July 1957, when he rejoined CWS. He worked out of Maple, Ontario, until his death, while on a field trip near Aultsville. While at Sackville, Mr. Boyer worked chiefly on waterfowl of the Nova Scotia-New Brunswick border region, with special emphasis on Pintails and Black Ducks. He also studied merganser- salmon interrelationships on the Miramichi River system, Woodcock, and the effects on bird popu lations of spruce budworm control spraying in the Upsalquitch area. -

Field Trips Guide Book for Photographers Revised 2008 a Publication of the Northern Virginia Alliance of Camera Clubs
Field Trips Guide Book for Photographers Revised 2008 A publication of the Northern Virginia Alliance of Camera Clubs Copyright 2008. All rights reserved. May not be reproduced or copied in any manner whatsoever. 1 Preface This field trips guide book has been written by Dave Carter and Ed Funk of the Northern Virginia Photographic Society, NVPS. Both are experienced and successful field trip organizers. Joseph Miller, NVPS, coordinated the printing and production of this guide book. In our view, field trips can provide an excellent opportunity for camera club members to find new subject matter to photograph, and perhaps even more important, to share with others the love of making pictures. Photography, after all, should be enjoyable. The pleasant experience of an outing together with other photographers in a picturesque setting can be stimulating as well as educational. It is difficullt to consistently arrange successful field trips, particularly if the club's membership is small. We hope this guide book will allow camera club members to become more active and involved in field trip activities. There are four camera clubs that make up the Northern Virginia Alliance of Camera Clubs McLean, Manassas-Warrenton, Northern Virginia and Vienna. All of these clubs are located within 45 minutes or less from each other. It is hoped that each club will be receptive to working together to plan and conduct field trip activities. There is an enormous amount of work to properly arrange and organize many field trips, and we encourage the field trips coordinator at each club to maintain close contact with the coordinators at the other clubs in the Alliance and to invite members of other clubs to join in the field trip. -

East Bay Hills Wind Project Mi'kmaq Ecological Knowledge Study
East Bay Hills Wind Project Mi’kmaq Ecological Knowledge Study Prepared for: Cape Breton Hydro Inc. December 2012 – Version 1 M.E.K.S. Project Team Jason Googoo, Project Manager Dave Moore, Author and Research Craig Hodder, Author and GIS Technician Mary Ellen Googoo, MEKS Interviewer John Sylliboy, MEKS Traditionalist Prepared by: Reviewed by: ___________________ ____________________ Craig Hodder, Author Jason Googoo, Manager Executive Summary This Mi’kmaq Ecological Knowledge Study, also commonly referred to as an MEKS or a Traditional Ecological Knowledge Study (TEKS), was developed by Membertou Geomatics Solutions (MGS) on behalf of Cape Breton Hydro Inc. (CBHI) for the proposed East Bay Hills Wind Power Project. This MEKS mandate is to consider land and water areas which the proposed project will utilize, and to identify what Mi’kmaq traditional use activities have occurred, or are currently occurring within, and what Mi’kmaq ecological knowledge presently exists in regards to the area. In order to ensure accountability and ethic responsibility of this MEKS, the MEKS development has adhered to the “Mi’kmaq Ecological Knowledge Protocol”. This protocol is a document that has been established by the Assembly of Nova Scotia Mi’kmaq Chiefs, which speaks to the process, procedures and results that are expected of a MEKS. The Mi’kmaq Ecological Knowledge Study consisted of two major components: • Mi’kmaq Traditional Land and Resource Use Activities , both past and present, • A Mi’kmaq Significance Species Analysis , considering the resources that are important to Mi’kmaq use. The Mi’kmaq Traditional Land and Resource Use Activities component utilized interviews as the key source of information regarding Mi’kmaq use in the Project Site and Study Area. -

Tectonic Setting of Middle Devonian to Lower Carboniferous Rocks in the Magdalen Basin P
Document generated on 10/06/2021 11:56 p.m. Atlantic Geology Tectonic setting of Middle Devonian to Lower Carboniferous rocks in the Magdalen Basin P. Durling and F. Marillier Volume 29, Number 3, November 1993 Article abstract Seismic reflection data in the Gulf of St. Lawrence were used to construct an URI: https://id.erudit.org/iderudit/ageo29_3rep03 isopach map of the Horton Group (Middle Devonian-Early Carboniferous) in the Magdalen Basin. The map shows that the Horton Group was deposited in See table of contents basins that developed parallel to the offshore extension of structural trends in New Brunswick and the Gaspe Peninsula. Horton Group strata were deposited in narrow linear fault bounded basins, presently up to 8 km deep, and in broad Publisher(s) sag basins up to 3 km deep. The sag basins may thin latterally to form sedimentary veneers less than 1 km thick. The fault bounded basins are mostly Atlantic Geoscience Society half-grabens and they developed during an early crustal extension phase. The largest and deepest basin strikes northeast-southwest, almost parallel to the ISSN Moncton Basin in New Brunswick. Sag basins and thin veneers are not fault controlled and are widespread, underlying most of the Magdalen Basin. They 0843-5561 (print) probably formed later than the fault bounded basins in response to the onset 1718-7885 (digital) of a regional subsidence phase. Deformation of Horton Group rocks in the Magdalen Basin is concentrated in Explore this journal fault zones up to 20 km wide, and mainly affected deep subbasins, where thrust faults and possible flower structures are observed. -
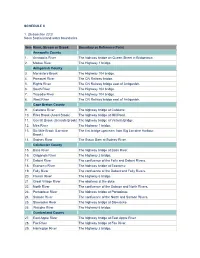
Nova Scotia Inland Water Boundaries Item River, Stream Or Brook
SCHEDULE II 1. (Subsection 2(1)) Nova Scotia inland water boundaries Item River, Stream or Brook Boundary or Reference Point Annapolis County 1. Annapolis River The highway bridge on Queen Street in Bridgetown. 2. Moose River The Highway 1 bridge. Antigonish County 3. Monastery Brook The Highway 104 bridge. 4. Pomquet River The CN Railway bridge. 5. Rights River The CN Railway bridge east of Antigonish. 6. South River The Highway 104 bridge. 7. Tracadie River The Highway 104 bridge. 8. West River The CN Railway bridge east of Antigonish. Cape Breton County 9. Catalone River The highway bridge at Catalone. 10. Fifes Brook (Aconi Brook) The highway bridge at Mill Pond. 11. Gerratt Brook (Gerards Brook) The highway bridge at Victoria Bridge. 12. Mira River The Highway 1 bridge. 13. Six Mile Brook (Lorraine The first bridge upstream from Big Lorraine Harbour. Brook) 14. Sydney River The Sysco Dam at Sydney River. Colchester County 15. Bass River The highway bridge at Bass River. 16. Chiganois River The Highway 2 bridge. 17. Debert River The confluence of the Folly and Debert Rivers. 18. Economy River The highway bridge at Economy. 19. Folly River The confluence of the Debert and Folly Rivers. 20. French River The Highway 6 bridge. 21. Great Village River The aboiteau at the dyke. 22. North River The confluence of the Salmon and North Rivers. 23. Portapique River The highway bridge at Portapique. 24. Salmon River The confluence of the North and Salmon Rivers. 25. Stewiacke River The highway bridge at Stewiacke. 26. Waughs River The Highway 6 bridge. -

South Western Nova Scotia
Netukulimk of Aquatic Natural Life “The N.C.N.S. Netukulimkewe’l Commission is the Natural Life Management Authority for the Large Community of Mi’kmaq /Aboriginal Peoples who continue to reside on Traditional Mi’Kmaq Territory in Nova Scotia undisplaced to Indian Act Reserves” P.O. Box 1320, Truro, N.S., B2N 5N2 Tel: 902-895-7050 Toll Free: 1-877-565-1752 2 Netukulimk of Aquatic Natural Life N.C.N.S. Netukulimkewe’l Commission Table of Contents: Page(s) The 1986 Proclamation by our late Mi’kmaq Grand Chief 4 The 1994 Commendation to all A.T.R.A. Netukli’tite’wk (Harvesters) 5 A Message From the N.C.N.S. Netukulimkewe’l Commission 6 Our Collective Rights Proclamation 7 A.T.R.A. Netukli’tite’wk (Harvester) Duties and Responsibilities 8-12 SCHEDULE I Responsible Netukulimkewe’l (Harvesting) Methods and Equipment 16 Dangers of Illegal Harvesting- Enjoy Safe Shellfish 17-19 Anglers Guide to Fishes Of Nova Scotia 20-21 SCHEDULE II Specific Species Exceptions 22 Mntmu’k, Saqskale’s, E’s and Nkata’laq (Oysters, Scallops, Clams and Mussels) 22 Maqtewe’kji’ka’w (Small Mouth Black Bass) 23 Elapaqnte’mat Ji’ka’w (Striped Bass) 24 Atoqwa’su (Trout), all types 25 Landlocked Plamu (Landlocked Salmon) 26 WenjiWape’k Mime’j (Atlantic Whitefish) 26 Lake Whitefish 26 Jakej (Lobster) 27 Other Species 33 Atlantic Plamu (Salmon) 34 Atlantic Plamu (Salmon) Netukulimk (Harvest) Zones, Seasons and Recommended Netukulimk (Harvest) Amounts: 55 SCHEDULE III Winter Lake Netukulimkewe’l (Harvesting) 56-62 Fishing and Water Safety 63 Protecting Our Community’s Aboriginal and Treaty Rights-Community 66-70 Dispositions and Appeals Regional Netukulimkewe’l Advisory Councils (R.N.A.C.’s) 74-75 Description of the 2018 N.C.N.S. -

Upgrading the Avon River Causeway During Highway 101 Twinning
Upgrading the Avon River Causeway During Highway 101 Twinning Dr. Bob Pett, NSTIR Environmental Services Alexander Wilson, CBCL Limited Transportation and Infrastructure Renewal Partner with NS Agriculture 9.5 km 6 lanes PEI Moncton NB Northumberland Strait Petitcodiac River NS Chignecto Bay Minas Basin Bay of Fundy Avon River Windsor Fundy Tides Salty- Silty Lake Pesaquid 1970 Fresh water Impacts on the Windsor Salt Marsh (Ramsar Wetland & IBA of Canada) Unlike the Petitcodiac – keeping an aboiteau EA completed in 2017 – currently working on design Project in planning for almost 20 years – including various environmental studies of the Avon River Estuary Contracted Acadia University, St. Mary’s University and CBWES Inc., between 2002 and 2018 to better understand the estuary and inform our design team to minimize impacts on salt marsh and mudflats. Baseline CRA Fisheries Study (Commercial, Recreational and Aboriginal) Contracted 3 partners for work between April 2017 and March 2019 ➢ Darren Porter, commercial fisher, ➢ Acadia University (Dr. Trevor Avery) ➢ Mi’kmaq Conservation Group Key study goal to better inform the detailed design team to improve fish passage through the aboiteau (sluice) Just before Christmas 2017, we engaged a team led by CBCL Limited to design an upgraded causeway and aboiteau system. Design Objectives Public Safety • Maintain corridor over Avon River for Highway 101 Twinning and continuity of rail, trail and utility services. • Continued protection of communities and agricultural land from the effects of flooding and sea level rise / climate change. Regulatory Requirements • Improve fish passage (EA Condition & Fisheries Act ). • Minimize environmental impacts (i.e., impact to salt marsh). • Consideration of potential negative impacts to asserted or established Mi’kmaq aboriginal or treaty rights. -

Northumberland Strait Crossing: Design Development of Precast Prestressed Bridge Structure
PRECAST CONCRETE SOLVES 100-YEAR-OLD PROBLEM Northumberland Strait Crossing: Design Development of Precast Prestressed Bridge Structure The authors describe the design development process of the $840 million (Canadian dollars) Northumberland Strait Crossing Project from conceptual design in 1987 to the final project design. The 13 km (8 mile) long bridge links Prince Edward Island with New Brunswick and mainland Canada. The current design is based on main bridge spans of 250 m (820 ft) to minimize the number of piers and foundations in the Strait. Each span consists of a continuous precast, prestressed concrete variable depth double cantilever girder Barry Lester, P.Eng. with a length of 190 m (623 ft) and a drop-in segment of 60 m President (197 ft). The design took into consideration unusually heavy SLG Stanley Consultants Inc. vehicle loads, high wind loads, seismic factors, very high Calgary, Alberta icepack forces and possible ship collisions. Precast concrete Canada production began in the summer of 1994 and the main spans will be erected beginning in September 1995. The project is scheduled for completion in the summer of 1997. he Northumberland Strait gotiating the Terms of Confederation, Crossing Project (NSCP) is a the Federal Government of Canada T 13 km (8 mile) long bridge (see promised to promote efficient commu Fig. 1), with associated approach nication between the island and the roads and shoreside facilities, joining mainland, a promise that has been ful Prince Edward Island to New filled by the payment of annual subsi Brunswick and mainland Canada (see dies to support the island ferry service Fig.