Multiproxy Reconstruction of Oceanographic Conditions in The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Preservation of Permian Fishes from the Marl Slate of County Durham
DEPARTMENT OF EARTH SCIENCES- DURHAM UNIVERSITY Prof. David Harper: [email protected] Dr Stuart Jones: [email protected] Preservation of Permian fishes from the Marl Slate of County Durham 1. Background The fossil fishes from the Permian Marl Slate (Fig.1) display superb preservation of palaeonis- A large fauna of beautifully-preserved fishes from coid fish from scales scattered sparingly to com- the Marl Slate was first described by William King plete whole specimens with extraordinary detail. in his landmark monograph on the Permian fossils of England, in 1850. The Marl Slate was deposited over large areas in middle Europe (extending from England to Russia) in a restricted, almost la- goonal environment; the setting is virtually unique in the stratigraphical record. Neverthe- less, we know little of the modes of preservation of the fishes, many of their life styles and their contribution to late Permian food webs. The po- tential too that soft parts may have been pre- served offers a unique opportunity to study the anatomy and decay processes of these Palaeozoic fishes. Outside Britain, the most closely compara- ble fish-bearing formation to the Marl Slate is the Kupferschiefer of Germany. This is a similar fine- grained, flaggy rock in which specimens are well preserved, flattened on individual laminae. The Kupferschiefer extends across north-west Europe (Lower Zechstein), and is regarded as marking a series of anoxic events prior to the main flooding of the Zechstein Basin in the first of five cycles Figure 1: Two examples of Palaeoniscus freislebenen- sis, Marl Slate, County Durham, UK The soft-part preservation remains because of se- lective preservation and subsequent diagenesis Key questions to study: to more resistant biopolymers. -
An Introduction to the Triassic: Current Insights Into the Regional Setting and Energy Resource Potential of NW Europe
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/322739556 An introduction to the Triassic: Current insights into the regional setting and energy resource potential of NW Europe Article in Geological Society London Special Publications · January 2018 DOI: 10.1144/SP469.1 CITATIONS READS 3 92 3 authors, including: Tom Mckie Ben Kilhams Shell U.K. Limited Shell Global 37 PUBLICATIONS 431 CITATIONS 11 PUBLICATIONS 61 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Reconstructing the Norwegian volcanic margin View project Paleocene of the Central North Sea: regional mapping from dense hydrocarbon industry datasets. View project All content following this page was uploaded by Ben Kilhams on 22 November 2019. The user has requested enhancement of the downloaded file. Downloaded from http://sp.lyellcollection.org/ by guest on January 26, 2018 An introduction to the Triassic: current insights into the regional setting and energy resource potential of NW Europe MARK GELUK1*, TOM MCKIE2 & BEN KILHAMS3 1Gerbrandylaan 18, 2314 EZ Leiden, The Netherlands 2Shell UK Exploration & Production, 1 Altens Farm Road, Nigg, Aberdeen AB12 3FY, UK 3Nederlandse Aardolie Maatschappij (NAM), PO Box 28000, 9400 HH Assen, The Netherlands *Correspondence: [email protected] Abstract: A review of recent Triassic research across the Southern Permian Basin area demon- strates the role that high-resolution stratigraphic correlation has in identifying the main controls on sedimentary facies and, subsequently, the distribution of hydrocarbon reservoirs. The depositio- nal and structural evolution of these sedimentary successions was the product of polyphase rifting controlled by antecedent structuration and halokinesis, fluctuating climate, and repeated marine flooding, leading to a wide range of reservoir types in a variety of structural configurations. -

Cretaceous-Tertiary Boundary Layer at Stevns Klint (Denmark): Copper and Copper(II) Porphyrins PAVLE I
J. Serb. Chem. Soc. 64(5 6)349-358(1999) UDC 547.979.732:546.56/.562 JSCS-2675 Original scientific paper Cretaceous-Tertiary boundary layer at Stevns Klint (Denmark): copper and copper(II) porphyrins PAVLE I. PREMOVI], NIKOLA D. NIKOLI], IVANA R. TONSA, DEJAN T. DULANOVI] and MIRJANA S. PAVLOVI]* Laboratory for Geochemistry and Cosmochemistry, Department of Chemistry, Faculty of Science, University of Ni{, P.O.Box 91, YU-18000 Ni{, Yugoslavia and *Vin~a Institute of Nuclear Sciences, P.O. Box 522, YU-11001 Belgrade, Yugoslavia (Received 2 September 1998, revised 11 February 1999) High concentrations (up to 4000 ppm) of copper(II) porphyrins have been detected in the kerogen of the Cretaceous/Tertiary (KT) boundary informal type sedi- ment, the Fish Clay, at Stevns Klint, Denmark. These pigments have also been found in the Danish KT sediment at Nye Kløv, which is about 300 km away from Stevns Klint. However, copper(II) porphyrins cannot be detected in the Danich boundary rock of the Dania site, which is separated from Stevns Klint by about 200 km. It is proposed that the kerogen copper(II) porphyrins are derived from humic materials of terrestrial (peat/soil) sources already enriched with these compounds, which were redeposited in the Danish KT boundary Basin. In addition, our results show that the kerogen copper(II) porphyrins are present, for comparison, in the Permian Kupferschiefer shale from Poland. The amounts are comparable to those in the Fish Clay kerogen. To our opinion, the same processes responsible for the kerogen copper(II) porphyrin enrichment have occurred in the Stevns Klint KT boundary sediment and in the Kupferschiefer. -

Coincidence of Photic Zone Euxinia and Impoverishment of Arthropods
www.nature.com/scientificreports OPEN Coincidence of photic zone euxinia and impoverishment of arthropods in the aftermath of the Frasnian- Famennian biotic crisis Krzysztof Broda1*, Leszek Marynowski2, Michał Rakociński1 & Michał Zatoń1 The lowermost Famennian deposits of the Kowala quarry (Holy Cross Mountains, Poland) are becoming famous for their rich fossil content such as their abundant phosphatized arthropod remains (mostly thylacocephalans). Here, for the frst time, palaeontological and geochemical data were integrated to document abundance and diversity patterns in the context of palaeoenvironmental changes. During deposition, the generally oxic to suboxic conditions were interrupted at least twice by the onset of photic zone euxinia (PZE). Previously, PZE was considered as essential in preserving phosphatised fossils from, e.g., the famous Gogo Formation, Australia. Here, we show, however, that during PZE, the abundance of arthropods drastically dropped. The phosphorous content during PZE was also very low in comparison to that from oxic-suboxic intervals where arthropods are the most abundant. As phosphorous is essential for phosphatisation but also tends to fux of the sediment during bottom water anoxia, we propose that the PZE in such a case does not promote the fossilisation of the arthropods but instead leads to their impoverishment and non-preservation. Thus, the PZE conditions with anoxic bottom waters cannot be presumed as universal for exceptional fossil preservation by phosphatisation, and caution must be paid when interpreting the fossil abundance on the background of redox conditions. 1 Euxinic conditions in aquatic environments are defned as the presence of H2S and absence of oxygen . If such conditions occur at the chemocline in the water column, where light is available, they are defned as photic zone euxinia (PZE). -
The Texan Permian and Its Mesozoic Types of Fossils.. .1891
WHITE, C.A. ...The Texan Permian and its Mesozoic types of fossils.. .1891. I*- b37(245 ¥58 jj DEPARTMENT OP THE INTERIOR OF THK UNITED STATES GEOLOGICAL SURVEY No. 77 THE TEXAN PERMIAN AND ITS MESOZOIC TYPES OF FOSSILS WASHINGTON GOVERNMENT PRINTING OFFICE 1891 LIBRARY CATALOGUE SLIPS. United States, Department of the interior. (U. S. geological survey). Department of the interior |- | Bulletin | of the | United States | geological survey | mo. 77 | [Seal of the department] | Washington \ government printing office | 1891 Second title: United States geological survey | J. W. Powell, director | | The Texau Permian | and its | Mesozoio types of fossils | by | Charles A. White | [Vignette] | Washington | government printing office | 1891 8°. 5lpp. 4 pi. White (Charles A.). United States geological survey | J.W. Powell, director | The Texau Permian | audits | Mesozoic types of fossils | by | Charles A. White | [Vignette] | Washington | government printing office | 1891. 8«. 5t pp. 4 pi. [UNITED STATES. Department of the interior. (U. S. geological survey). Bulletin 77.] United States geological survey J. W. Powell, director | | The Texan Permian | and its | Mesozoic types of fossils | by | Charles A. White | [Vignette] | Washingto:a | government printing office | 1891 8". 5lpp. 4 pi. [UNITED STATES. Department <tf the interior. (U. S. geological survey). Bnlletin 77.] A.D'VBRTISEMKN'T. [Bulletin No. 77.] The publications of the United States Geological Survey are issued in accordance with the statute approved March 3, 1879, which declares that "Thepublicationsof tb.eGeological Survey shall consist of the annual report of operations, geological and economic maps illustrating the resources and classification of the lauds, and reports upon general and economic geology and paleontology. -

Geology and Paragenesis of the Boseto Copper Deposits, Kalahari
GEOLOGY AND PARAGENESIS OF THE BOSETO COPPER DEPOSITS, KALAHARI COPPERBELT, NORTHWEST BOTSWANA by Wesley S. Hall A thesis submitted to the Faculty and the Board of Trustees of the Colorado School of Mines in partial fulfillment of the requirements for the degree of Master of Science (Geology) Golden, Colorado Date _________________________ Signed: ______________________________ Wesley S. Hall Signed: ______________________________ Dr. Murray W. Hitzman Thesis Advisor Golden, Colorado Date _________________________ Signed: ______________________________ Dr. John D. Humphrey Associate Professor and Head Department of Geology & Geological Engineering ii ABSTRACT Detailed lithostratigraphic, structural, and petrographic studies coupled with fluid inclusion and stable isotopic analyses and geochronological studies indicate that the Boseto copper deposits formed initially during diagenesis as metalliferous brines ascended along basin faults and moved along a stratigraphic redox boundary between continental red beds and an overlying reduced marine siliciclastic sequence. The hanging wall rocks to copper-silver ore zones comprise comprises a series of at least three stacked coarsening upwards cycles deposited in a deltaic depositional setting. Early copper mineralization may have been accompanied by regionally extensive albitization. Later multiple pulses of faulting and hydrothermal fluid flow associated with a southeast-vergent folding event in the Ghanzi-Chobe belt resulted in extensive networks of bedding-parallel and discordant quartz-carbonate-(Cu-Fe-sulfide) veins. This contractional deformation-related vein and shear system was responsible for significant remobilization of pre-existing vertically and laterally zoned copper sulfide minerals into high- grade zones by hot (250-300˚C), syn-orogenic, metamorphic-derived hydrothermal fluids. Orientation analysis indicates that the mineralized veins probably formed in association with a flexural slip folding processes. -

Limestone Landscapes: a Geodiversity Audit and Action Plan for The
Limestone Landscapes - a geodiversity audit and action plan for the Durham Magnesian Limestone Plateau Geology and Landscape England Programme Open Report OR/09/007 BRITISH GEOLOGICAL SURVEY GEOLOGY AND LANDSCAPE ENGLAND PROGRAMME OPEN REPORT OR/09/007 Limestone Landscapes - a geodiversity audit and action The National Grid and other Ordnance Survey data are used plan for the Durham Magnesian with the permission of the Con- troller of Her Majesty’s Station- ery Office. Limestone Plateau Licence No: 100017897/ 2009. Keywords geodiversity, Durham, Permian, D J D Lawrence Limestone, Landscape. National Grid Reference Editor SW corner 429800,521000 Centre point 438000,544000 A H Cooper NE corner 453400,568000 Front cover The Magnesian Limestone at Marsden Bay Bibliographical reference LAWRENCE, D J D. 2009. Limestone Landscapes - a geodiversity audit and action plan for the Durham Magnesian Limestone Plateau. British Geological Survey Open Report, OR/09/007. 114pp. Copyright in materials derived from the British Geological Survey’s work is owned by the Natural Environment Research Council (NERC) and/or the authority that commissioned the work. You may not copy or adapt this publication without first obtaining permission. Contact the BGS Intellectual Property Rights Section, British Geological Sur- vey, Keyworth, E-mail [email protected]. You may quote extracts of a reasonable length without prior permission, provided a full acknowledgement is given of the source of the extract. Maps and diagrams in this book use topography based on Ord- nance -

Eidesstattliche Erklärung
Eidesstattliche Erklärung Vor- und Zuname: Martin Stockhausen Geburtsdatum: 15.09.1979 Geburtsort: Solingen Hiermit erkläre ich an Eides statt, dass ich die vorgelegte Dissertation selbständig und ohne unzulässige fremde Hilfe angefertigt und verfasst habe, dass alle Hilfsmittel und sonstigen Hilfen angegeben und dass alle Stellen, die ich wörtlich oder dem Sinne nach aus anderen Veröffentlichungen entnommen habe, kenntlich gemacht worden sind; dass die Dissertation ganz oder teilweise noch nicht zu einem früheren Zeitpunkt an der Christian-Albrechts- Universität zu Kiel oder einer anderen in- oder ausländischen Hochschule als Dissertation eingereicht worden ist. Die Dissertation entstand unter Einhaltung der Regeln guter wissenschaftlicher Praxis der Deutschen Forschungsgemeinschaft. Kiel, 24.09.2015 gez. Martin Stockhausen EXPERIMENTAL SIMULATION OF HYDROCARBON EXPULSION Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät der Christian-Albrechts-Universität zu Kiel vorgelegt von Martin Stockhausen Kiel, 2015 Erster Gutachter: Prof. Dr. Lorenz Schwark Zweiter Gutachter: Prof. Dr. Brain Horsfield Tag der mündlichen Prüfung: 30.11.2015 Zum Druck genehmigt: 30.11.2015 gez. Prof. Dr. Wolfgang J. Duschl, Dekan Abstract Investigating generation, expulsion and primary migration usually suffers from inadequate methodologies, failing in the provision of natural conditions, prevailing during the genesis of oil and gas. Destructive sample preparation, inappropriate pressure regimes and pyrolysis in closed mode and/or in the absence of water caused results not representative for natural processes. To overcome these limitations, a newly designed apparatus was developed and built, capable to perform pyrolysis experiments with intact rock samples under pressure regimes, prevailing during catagenesis. In detail, lithostatic (or overburden-) pressures and hydrostatic (or pore-) pressures, corresponding to 3000 m depth and beyond, can be simulated by this apparatus, the “Expulsinator” device. -

Hydrocarbon Plays from West Poland: Zechstein Limestone and Main Dolomite
Hydrocarbon Plays from West Poland: Zechstein Limestone and Main Dolomite Paweł Zdanowski & Tomasz Solarski PGNiG SA (Polish Oli & Gas Company), Exploration and Production Branch Underexplored Plays - Part III 31-X-2018 & 01-XI-2018 Stavanger Hydrocarbon Plays from West Poland: Zechstein Limestone (Ca1) and Main Dolomite (Ca2) Presentation outline: Palaeogeography of Zechstein • Southern Permian Basin and Northern Permian Basin • Correlation of carbonate units between SBP and NPB Zechstein Limestone (Brońsko Gas Field) • Hydrocarbon Play. • Pattern Recognition from Seismic. Main Dolomite (BMB and LMG oil & gas fields) • Hydrocarbon Play. • Pattern Recognition from Seismic. • Analysis of Seismic Attributes. • Seismic Modeling. Late Permian Paleogeography from Blakely (2014) Tucker, 2016 After Scotese, 2002 European Southern Permian Basin Zechstein Sea Connected to Pantalassa Ocean 2000km to N/NE. Possible connection to Paleo-Tethys through the Polish Sub-Basin to the SE ?? Palaeo-latitude: 10-20°. Climate extremely arid. After Scotese, 2012 Sketch map of Permian sedimentary basin in north-west Europe Dziękuję za uwagę Stavanger Warsaw Millennium Atlas: Petroleum Geology of the Central and Northern North Sea, 2003 Polish Zechstein Basin ….stratigraphy Leine Na2 A2 Stassfurt Ca2 A1g Na1 A1d Werra Ca1 Wagner & Peryt. 1997 Słowakiewicz & Mikołajewski, 2009 Tucker, 2016 Comparison of the Zechstein development of the NPB with the classic SPB Evans et al. 2003 Madeleine et al. 2018 Zechstein Limestone – Ca1 (≈ Argyll Carbonate Member) Zechstein basin, showing the position of the Brandenburg-Wolsztyn-Pogorzela palaeo-High Basin facies of the Zechstein Limestone in Poland, showing the occurrence of isolated reefs related to the Brandenburg-Wolsztyn-Pogorzela palaeo-High. isolated reefs (rimmed platform) Peryt et al. -

3.2 Permian but More Differentiated Rocks Occur in the P-1 and W-1 Wells
28 Boundaries: The lower boundary is primarily marked tance early in Permian. The initial phase of subsidence by the sonic velocity which is high and uniform in the was accompanied by extensive subaerial volcanism. underlying metamorphic greenstone, and relatively This was followed by a period of oxidation and erosion lower and nervous in the present series. The upper under desert conditions and deposition of red beds and boundary is between the present mainly greyish series sabkha sediments in the two Permian basins (fig. 19). and the overlying reddish series of assumed Rot These rocks are included in the Rotliegendes Group as liegendes age. The log motifs are not significant, but originally established by Werner (1786). Continuous the change from high gamma ray readings above to subsidence and transgression of the sea, but with a relatively lower below is used for location of the restricted connection to the ocean, lead to the forma boundary. tion of the evaporites of the Zechstein Group. Distribution: The Early Carboniferous deposits are probably present in major parts of the Danish Central Rotliegendes Group Graben, but due to lack of data (seismic mapping is Autunian critical), a delineation of the extension is not possible. Type section: For the present being the P-1 well, Geological age: Based on studies of the miospores, 10541-10865' b.KB may suffice as a reference for the part of the series is dated to Early Carboniferous, Late Rotliegendes volcanics and underlying sediments be Visean or Early Namurian (Bertelsen 1978). - The longing to the Autunian stage. However, it should be lower part of the series has not been dated, thus it may noted that this correlation is based purely on be of Carboniferous or Devonian age. -
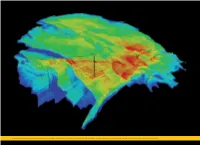
3D View of the Top Rotliegend Reservoir Showing the Depth Structure of the Groningen Gasfield and the Slochteren-1 Exploration Well
SPBA-Compleet 22-04-10 14:24 Pagina 270 3D view of the top Rotliegend reservoir showing the depth structure of the Groningen gasfield and the Slochteren-1 exploration well. The perspective is from the SSW. Field length and width are approximately 40 km and 30 km, respectively. Colour scale: red is at 2600 m and dark blue is at 3000 m below mean sea level. SPBA-Compleet 22-04-10 14:24 Pagina 271 Chapter 15 — Reserves and production history Chapter 15 Reserves and production history Authors Bibliographic reference Jaap Breunese (TNO), Jan Andersen (DEA), Sven Brinkman (BGR), Paweł Jagosiak (POGC), Steffen Bjørn Olsen (DEA), Tadeusz Peryt (PGI), Joachim Piske (Consultant), Pawel Poprawa (PGI), Breunese, J.N., Andersen, J.H., Brinkman, S., Jagosiak, P., Karnin, W-D., Karnkowski, P.H., Kombrink, H., Messner, J., Wolf-Dieter Karnin (Consultant), Paweł Karnkowski (University of Warsaw), Henk Kombrink (TNO), Jan Roelofsen (IHS Energy), Susan Stoker (BGS), Nigel Smith (BGS), Geoff Swann (UK Department of Energy Mijnlieff, H., Olsen, S.B., Peryt, T.M., Piske, J., Poprawa, P., Roelofsen, J.W., Stoker, S.J., Smith, N.J.P., Swann, G., Juergen Messner (State Authority for Mining, Energy and Geology, Germany), Harmen Mijnlieff (TNO), and Climate Change), Maria Waksmundzka (PGI) and Hans Veldkamp (TNO) Waksmundzka, M.I. & Veldkamp, J.G., 2010. Reserves and production history. In: Doornenbal, J.C. and Stevenson, A.G. (editors): Petroleum Geological Atlas of the Southern Permian Basin Area. EAGE Publications b.v. (Houten): 271-281. 0¡ 5¡E 10¡E 15¡E 20¡E 1 Introduction Fields related to Paleozoic source rocks Cenozoic Figures 15.1 and 15.2 show all 1392 oil and gas accumulations (grouped into 1244 fields) discovered so Cretaceous far within the SPB area. -

Geology of the Groningen Field – an Overview
Netherlands Journal of Geosciences — Geologie en Mijnbouw |96 – 5 | s3–s15 | 2017 doi:10.1017/njg.2017.22 Geology of the Groningen field – an overview JandeJager1,∗ & Clemens Visser2 1 Nassaukade 42, 2281 XD Rijswijk, the Netherlands 2 Nederlandse Aardolie Maatschappij B.V., Schepersmaat 2, 9405 TA Assen, the Netherlands ∗ Corresponding author. Email: [email protected] Manuscript received: 27 February 2017, accepted: 21 July 2017 Abstract After more than half a century of production and with some 350 wells, the Groningen gas field must be one of the best-studied gas fields in the world. Initially, it was considered to be relatively simple and behaving like one big tank. Now that it is entering a phase of declining production it has become clear that many subtleties are not fully understood yet. Prediction and management of subsidence and induced earth tremors require a detailed understanding of the field geology. In addition, an optimum gas recovery is only possible if details of, for example, reservoir quality distribution and faulting, that did not appear relevant before, are well understood. The large Groningen field comprises a structurally high block during much of its history, probably already from Devonian times onwards. The desert sandstones of the Rotliegend reservoir exhibit a strong south-to-north proximal–distal relationship. Whilst diagenesis has in many fields led to deterioration of reservoir properties, this effect is small in the Groningen field. The field is dipping to the north, and bounded by a series ofnormal faults in the west, south and east. Almost all faults are normal extensional faults, but locally inverse reactivation has led to small pop-up structures.