N Vitro Classical Turtles Conditioning of Abducens Nerve Discharge In
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Pharnygeal Arch Set - Motor USMLE, Limited Edition > Neuroscience > Neuroscience
CNs 5, 7, 9, 10 - Pharnygeal Arch Set - Motor USMLE, Limited Edition > Neuroscience > Neuroscience PHARYNGEAL ARCH SET, CNS 5, 7, 9, 10 • They are derived from the pharyngeal (aka branchial) arches • They have special motor and autonomic motor functions CRANIAL NERVES EXIT FROM THE BRAINSTEM CN 5, the trigeminal nerve exits the mid/lower pons.* CN 7, the facial nerve exits the pontomedullary junction.* CN 9, the glossopharyngeal nerve exits the lateral medulla.* CN 10, the vagus nerve exits the lateral medulla.* CRANIAL NERVE NUCLEI AT BRAINSTEM LEVELS Midbrain • The motor trigeminal nucleus of CN 5. Nerve Path: • The motor division of the trigeminal nerve passes laterally to enter cerebellopontine angle cistern. Pons • The facial nucleus of CN 7. • The superior salivatory nucleus of CN 7. Nerve Path: • CN 7 sweeps over the abducens nucleus as it exits the brainstem laterally in an internal genu, which generates a small bump in the floor of the fourth ventricle: the facial colliculus • Fibers emanate from the superior salivatory nucleus, as well. Medulla • The dorsal motor nucleus of the vagus, CN 10 • The inferior salivatory nucleus, CN 9 1 / 3 • The nucleus ambiguus, CNs 9 and 10. Nerve Paths: • CNs 9 and 10 exit the medulla laterally through the post-olivary sulcus to enter the cerebellomedullary cistern. THE TRIGEMINAL NERVE, CN 5  • The motor division of the trigeminal nerve innervates the muscles of mastication • It passes ventrolaterally through the cerebellopontine angle cistern and exits through foramen ovale as part of the mandibular division (CN 5[3]). Clinical Correlation - Trigeminal Neuropathy THE FACIAL NERVE, CN 7  • The facial nucleus innervates the muscles of facial expression • It spans from the lower pons to the pontomedullary junction. -

Projections from the Trigeminal Nuclear Complex to the Cochlear Nuclei: a Retrograde and Anterograde Tracing Study in the Guinea Pig
Journal of Neuroscience Research 78:901–907 (2004) Projections From the Trigeminal Nuclear Complex to the Cochlear Nuclei: a Retrograde and Anterograde Tracing Study in the Guinea Pig Jianxun Zhou and Susan Shore* Department of Otolaryngology and Kresge Hearing Research Institute, University of Michigan, Ann Arbor, Michigan In addition to input from auditory centers, the cochlear cuneate nucleus innervation of cochlear nucleus has been nucleus (CN) receives inputs from nonauditory centers, hypothesized to convey information about head and pinna including the trigeminal sensory complex. The detailed position for the purpose of localizing a sound source in anatomy, however, and the functional implications of the space (Young et al., 1995). In addition, interactions be- nonauditory innervation of the auditory system are not tween somatosensory and auditory systems have been fully understood. We demonstrated previously that the linked increasingly to phantom sound perception, also trigeminal ganglion projects to CN, with terminal labeling known as tinnitus. This is demonstrated in the observa- most dense in the marginal cell area and secondarily in tions that injuries of the head and neck region can lead to the magnocellular area of the ventral cochlear nucleus the onset of tinnitus in patients with no hearing loss (VCN). We continue this line of study by investigating the (Lockwood et al., 1998). projection from the spinal trigeminal nucleus to CN in We demonstrated previously projections from the guinea pig. After injections of the retrograde tracers Flu- trigeminal ganglion to CN in guinea pigs (Shore et al., oroGold or biotinylated dextran amine (BDA) in VCN, 2000). Terminal labeling of trigeminal ganglion projec- labeled cells were found in the spinal trigeminal nuclei, tions to the CN was found to be most dense in the most densely in the pars interpolaris and pars caudalis marginal cell area and secondarily in the magnocellular with ipsilateral dominance. -

Direct Projections from Cochlear Nuclear Complex to Auditory Thalamus in the Rat
The Journal of Neuroscience, December 15, 2002, 22(24):10891–10897 Direct Projections from Cochlear Nuclear Complex to Auditory Thalamus in the Rat Manuel S. Malmierca,1 Miguel A. Mercha´n,1 Craig K. Henkel,2 and Douglas L. Oliver3 1Laboratory for the Neurobiology of Hearing, Institute for Neuroscience of Castilla y Leo´ n and Faculty of Medicine, University of Salamanca, 37007 Salamanca, Spain, 2Wake Forest University School of Medicine, Department of Neurobiology and Anatomy, Winston-Salem, North Carolina 27157-1010, and 3University of Connecticut Health Center, Department of Neuroscience, Farmington, Connecticut 06030-3401 It is known that the dorsal cochlear nucleus and medial genic- inferior colliculus and are widely distributed within the medial ulate body in the auditory system receive significant inputs from division of the medial geniculate, suggesting that the projection somatosensory and visual–motor sources, but the purpose of is not topographic. As a nonlemniscal auditory pathway that such inputs is not totally understood. Moreover, a direct con- parallels the conventional auditory lemniscal pathway, its func- nection of these structures has not been demonstrated, be- tions may be distinct from the perception of sound. Because cause it is generally accepted that the inferior colliculus is an this pathway links the parts of the auditory system with prom- obligatory relay for all ascending input. In the present study, we inent nonauditory, multimodal inputs, it may form a neural have used auditory neurophysiology, double labeling with an- network through which nonauditory sensory and visual–motor terograde tracers, and retrograde tracers to investigate the systems may modulate auditory information processing. -

Auditory and Vestibular Systems Objective • to Learn the Functional
Auditory and Vestibular Systems Objective • To learn the functional organization of the auditory and vestibular systems • To understand how one can use changes in auditory function following injury to localize the site of a lesion • To begin to learn the vestibular pathways, as a prelude to studying motor pathways controlling balance in a later lab. Ch 7 Key Figs: 7-1; 7-2; 7-4; 7-5 Clinical Case #2 Hearing loss and dizziness; CC4-1 Self evaluation • Be able to identify all structures listed in key terms and describe briefly their principal functions • Use neuroanatomy on the web to test your understanding ************************************************************************************** List of media F-5 Vestibular efferent connections The first order neurons of the vestibular system are bipolar cells whose cell bodies are located in the vestibular ganglion in the internal ear (NTA Fig. 7-3). The distal processes of these cells contact the receptor hair cells located within the ampulae of the semicircular canals and the utricle and saccule. The central processes of the bipolar cells constitute the vestibular portion of the vestibulocochlear (VIIIth cranial) nerve. Most of these primary vestibular afferents enter the ipsilateral brain stem inferior to the inferior cerebellar peduncle to terminate in the vestibular nuclear complex, which is located in the medulla and caudal pons. The vestibular nuclear complex (NTA Figs, 7-2, 7-3), which lies in the floor of the fourth ventricle, contains four nuclei: 1) the superior vestibular nucleus; 2) the inferior vestibular nucleus; 3) the lateral vestibular nucleus; and 4) the medial vestibular nucleus. Vestibular nuclei give rise to secondary fibers that project to the cerebellum, certain motor cranial nerve nuclei, the reticular formation, all spinal levels, and the thalamus. -

Use of Calcium-Binding Proteins to Map Inputs in Vestibular Nuclei of the Gerbil
THE JOURNAL OF COMPARATIVE NEUROLOGY 386:317–327 (1997) Use of Calcium-Binding Proteins to Map Inputs in Vestibular Nuclei of the Gerbil GOLDA ANNE KEVETTER* AND ROBERT B. LEONARD Departments of Otolaryngology, Anatomy and Neurosciences, and Physiology and Biophysics, University of Texas Medical Branch, Galveston, Texas 77555 ABSTRACT We wished to determine whether calbindin and/or calretinin are appropriate markers for vestibular afferents, a population of neurons in the vestibular nuclear complex, or cerebellar Purkinje inputs. To accomplish this goal, immunocytochemical staining was observed in gerbils after lesions of the vestibular nerve central to the ganglion, the cerebellum, or both. Eleven to fourteen days after recovery, the brain was processed for immunocytochemical identification of calretinin and calbindin. After lesion of the vestibular nerve, no calretinin staining was seen in any of the vestibular nuclei except for a population of intrinsic neurons, which showed no obvious change in number or staining pattern. Calbindin staining was reduced in all nuclei except the dorsal part of the lateral vestibular nuclei. The density of staining of each marker, measured in the magnocellular medial vestibular nucleus, was signifi- cantly reduced. After the cerebellar lesion, no differences in calretinin staining were noted. However, calbindin staining was greatly reduced in all nuclei. The density of staining, measured in the caudal medial vestibular nucleus, was significantly lower. After a combined lesion of the cerebellum and vestibular nerve, the distribution and density of calretinin staining resembled that after vestibular nerve section alone, whereas calbindin staining was no longer seen. This study demonstrates that calretinin and calbindin are effective markers for the identification of vestibular afferents. -
Auditory Nerve.Pdf
1 Sound waves from the auditory environment all combine in the ear canal to form a complex waveform. This waveform is deconstructed by the cochlea with respect to time, loudness, and frequency and neural signals representing these features are carried into the brain by the auditory nerve. It is thought that features of the sounds are processed centrally along parallel and hierarchical pathways where eventually percepts of the sounds are organized. 2 In mammals, the neural representation of acoustic information enters the brain by way of the auditory nerve. The auditory nerve terminates in the cochlear nucleus, and the cochlear nucleus in turn gives rise to multiple output projections that form separate but parallel limbs of the ascending auditory pathways. How the brain normally processes acoustic information will be heavily dependent upon the organization of auditory nerve input to the cochlear nucleus and on the nature of the different neural circuits that are established at this early stage. 3 This histology slide of a cat cochlea (right) illustrates the sensory receptors, the auditory nerve, and its target the cochlear nucleus. The orientation of the cut is illustrated by the pink line in the drawing of the cat head (left). We learned about the relationship between these structures by inserting a dye-filled micropipette into the auditory nerve and making small injections of the dye. After histological processing, stained single fibers were reconstruct back to their origin, and traced centrally to determine how they terminated in the brain. We will review the components of the nerve with respect to composition, innervation of the receptors, cell body morphology, myelination, and central terminations. -

Acoustically Responsive Fibers in the Vestibular Nerve of the Cat
The Journal of Neuroscience, October 1994, 74(10): 6056-6070 Acoustically Responsive Fibers in the Vestibular Nerve of the Cat Michael P. McCue1v2*a and John J. Guinan, Jr.r.2.3-4 ‘Eaton-Peabody Laboratory of Auditory Physiology, Department of Otolaryngology, Massachusetts Eye and Ear Infirmary, Boston, Massachusetts 02114, 2Harvard-MIT Division of Health Science and Technology and Research Laboratory of Electronics, and 3Department of Electrical Engineering and Computer Science, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139, and 4Department of Otology and Laryngology, Harvard Medical School, Boston, Massachusetts 02115 Recordings were made from single afferent fibers in the and levels within the normal range of human hearing. We inferior vestibular nerve, which innervates the saccule and suggest a number of auditory roles that these fibers may posterior semicircular canal. A substantial portion of the fi- play in the everyday life of mammals. bers with irregular background activity increased their firing [Key words: saccule, otoliths, auditory system, mamma- in response to moderately intense clicks and tones. lian sound reception, middle-ear muscles, cochlear nucleus] In responsive fibers, acoustic clicks evoked action poten- tials with minimum latencies of I 1 .O msec. Fibers fell into The vertebrate inner ear contains several senseorgans involved two classes, with the shortest latency either to condensation in the maintenance of equilibrium and the detection of vibra- clicks (PUSH fibers) or to rarefaction clicks (PULL fibers). tion. The precise sensory role assumedby homologous organs Low-frequency (800 Hz) tone bursts at moderately high sound varies among species.For example, the sacculeis thought to act levels (>80 dB SPL) caused synchronization of spikes to asa linear accelerometerin mammals(Fernindez and Goldberg, preferred phases of the tone cycle. -

Processing in the Cochlear Nucleus
Processing in The Cochlear Nucleus Alan R. Palmer Medical Research Council Institute of Hearing Research University Park Nottingham NG7 2RD, UK The Auditory Nervous System Cortex Cortex MGB Medial Geniculate Body Excitatory GABAergic IC Inferior Colliculus Glycinergic DNLL Nuclei of the Lateral Lemniscus Lateral Lemniscus Cochlear Nucleus DCN PVCN MSO Lateral Superior Olive AVCN Medial Superior Olive Cochlea MNTB Medial Nucleus of the Trapezoid Body Superior Olive The cochlear nucleus is the site of termination of fibres of the auditory nerve Cochlear Nucleus Auditory Nerve Cochlea 1 Frequency Tonotopicity Basilar membrane Inner hair cell Auditory nerve Fibre To the brain Each auditory-nerve fibre responds only to a narrow range of frequencies Tuning curve Action potential Evans 1975 2 Palmer and Evans 1975 There are many overlapping single-fibre tuning curves in the auditory nerve Audiogram Palmer and Evans 1975 Tonotopic Organisation Lorente - 1933 3 Tonotopic Organisation Base Anterior Cochlea Characteristic Basilar Membrane Frequency Hair Cells Auditory Nerve Apex Cochlear Nucleus Spiral Ganglion Posterior Tonotopic projection of auditory-nerve fibers into the cochlear nucleus Ryugo and Parks, 2003 The cochlear nucleus: the first auditory nucleus in the CNS Best frequency Position along electrode track (mm) Evans 1975 4 stellate (DCN) Inhibitory Synapse Excitatory Synapse DAS to inferior colliculus cartwheel fusiform SUPERIOR OLIVARY giant COMPLEX INFERIOR COLLICULUS granule vertical vertical OCB AANN to CN & IC via TB golgi DORSAL -

Supplemental Figure 1. Egr2 and Hoxb1 Are Expressed in and Delete
Supplemental Figure 1. Egr2Cre and Hoxb1Cre are expressed in and delete Atoh1flox from different regions of the developing neural tube. (A, A’) E9.5 Egr2Cre; ROSAR26R embryo wholemount stained with Xgal. Cre expression drives β-galactosidase expression only in rhombomeres 3 and 5. White and black dotted lines here and in panels E and E’ denote the anterior and posterior boundaries of rhombomere 4, respectively. (B, C) Coronal sections from a P30 Egr2Cre; ROSAR26R animal showing β-galactosidase expression in the CN. Large cells and granule cells are labeled in the AVCN and DCN, but to a much lesser extent in the PVCN. (D) Xgal staining in coronal sections of the AAN of the same animal. High levels of expression are seen throughout the AAN, demonstrating that these nuclei arise predominantly from rhombomeres 3 and 5. (E, E’) E9.5 Hoxb1Cre; ROSAR26R embryo wholemount stained with Xgal. Cre expression drives β-galactosidase expression posterior to the anterior boundary of rhombomere 4. (F, G) Coronal sections from a P30 Hoxb1Cre; ROSAR26R animal showing heavy expression of β-galactosidase in the PVCN and DCN with much lower levels of expression in the AVCN. (H) Xgal staining in coronal sections of the AAN of the same Hoxb1Cre; ROSAR26R animal. High levels of expression are seen throughout the AAN with the exception of the MNTB, where only some of the neurons are labeled. (I-N) Atoh1 in situ hybridization in coronal hindbrain sections from E15.5 wildtype (I, L), Egr2Cre; Atoh1CKO (J, M) and Hoxb1; Atoh1CKO (K, N) embryos. Red arrows show the absence of Atoh1 mRNA in the neuroepithelium that contributes neurons to the developing AES, AVCN, cRL, and DCN, while expression of the gene is maintained in other regions where Cre is not expressed. -

Physiology of the Cochlear Nucleus
Physiology of the cochlear nucleus Spherical vs globular bushy cells Circuitry in dorsal cochlear nucleus Differentiating cell types in the cochlear nucleus • By anatomical shape • Using soma shape with Nissl stains: Osen (spherical, globular, multipolar, pyramidal, giant, granule) • Using dendritic structure in Golgi stains: Bushy, stellate, octopus, cartwheel • Using their axonal projection patterns • Imaginative names help (unipolar brush cells, chestnut cells, etc.) • By physiological response type (PST shape) • PSTH shape: Primary-like, PL with notch, chopper, pauser, build-up, onset • Response area characteristics (type I, II, III, IV) Brad May, JHU Correlating anatomy with physiology (form & function) has been a persistent theme of CN research Notch PLn Brad May, JHU Variety of PSTs seen in the CN Adams, 1976 Joseph C. Adams M.I.T. Brain slices of CN show bushy and stellate cells have different membrane properties Wu and Oertel, 1984 Intracellular recording and staining is definitive, but… Combined intracellular recording (to get PSTs) with staining allows structure/function relations to be studied. However, neither the cell type nor the PST type are absolute indicators. To IC (bilateral) • All principal cells receive ANF input • Note that with the exception of the bushy cells, all other cell types receive additional inputs that would DCN be expected to make them To SOC more selective than ANFs • On the other hand there are synapses on the ‘bush’ of the BC but little is known about them so they are ignored. • How would you determine -
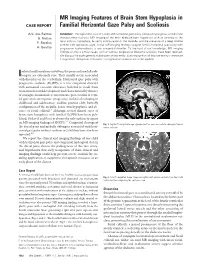
MR Imaging Features of Brain Stem Hypoplasia in Familial Horizontal
MR Imaging Features of Brain Stem Hypoplasia in CASE REPORT Familial Horizontal Gaze Palsy and Scoliosis A.V. dos Santos SUMMARY: We report the case of a child with horizontal gaze palsy, pendular nystagmus, and discrete S. Matias thoracolumbar scoliosis. MR imaging of the brain depicted pons hypoplasia with an absence of the facial colliculi, hypoplasia, butterfly configuration of the medulla, and the presence of a deep midline P. Saraiva pontine cleft (split pons sign). These MR imaging findings suggest familial horizontal gaze palsy with A. Goula˜o progressive kyphoscoliosis, a rare congenital disorder. To the best of our knowledge, MR imaging findings of only 4 similar cases, with or without progressive idiopathic scoliosis, have been reported. We discuss the pathogenesis substratum of this entity. Early recognition of this rare entity is important if supportive therapeutic measures in progressive scoliosis are to be applied. solated malformations involving the pons and medulla ob- Ilongata are extremely rare. They usually occur associated with disorders of the cerebellum. Horizontal gaze palsy with progressive scoliosis (HGPPS) is a rare congenital disorder with autosomal recessive inherence, believed to result from cranial nuclear maldevelopment and characterized by absence of conjugate horizontal eye movements, preservation of verti- cal gaze and convergence, progressive scoliosis developing in childhood and adolescence, midline pontine cleft, butterfly configuration of the medulla, brain stem hypoplasia, and ab- sence of facial colliculi.1 Although several clinical cases of brain stem hypoplasia with familial HGPPS have been pub- lished, Pieh et al and Rossi et al were the only authors to report on MR imaging findings of HGPPS.2,3 Congenital cleavage of Fig 1. -

Trigeminal Ganglion Innervates the Auditory Brainstem
THE JOURNAL OF COMPARATIVE NEUROLOGY 419:271–285 (2000) Trigeminal Ganglion Innervates the Auditory Brainstem SUSAN E. SHORE,1,2* ZOLTAN VASS,3 NOEL L. WYS,1 AND RICHARD A. ALTSCHULER1 1Kresge Hearing Research Institute, University of Michigan, Ann Arbor, Michigan 48109-0506 2Department of Otolaryngology, Medical College of Ohio, Toledo, Ohio 43699 3Department of Otolaryngology, Albert Szent-Gyorgyi Medical University, Szeged, Hungary ABSTRACT A neural connection between the trigeminal ganglion and the auditory brainstem was investigated by using retrograde and anterograde tract tracing methods: iontophoretic injec- tions of biocytin or biotinylated dextran-amine (BDA) were made into the guinea pig trigem- inal ganglion, and anterograde labeling was examined in the cochlear nucleus and superior olivary complex. Terminal labeling after biocytin and BDA injections into the ganglion was found to be most dense in the marginal cell area and secondarily in the magnocellular area of the ventral cochlear nucleus (VCN). Anterograde and retrograde labeling was also seen in the shell regions of the lateral superior olivary complex and in periolivary regions. The labeling was seen in the neuropil, on neuronal somata, and in regions surrounding blood vessels. Retrograde labeling was investigated using either wheatgerm agglutinin- horseradish peroxidase (WGA-HRP), BDA, or a fluorescent tracer, iontophoretically injected into the VCN. Cells filled by retrograde labeling were found in the ophthalmic and mandib- ular divisions of the trigeminal ganglion. We have previously shown that these divisions project to the cochlea and middle ear, respectively. This study provides the first evidence that the trigeminal ganglion innervates the cochlear nucleus and superior olivary complex.