The Phenotypic Morphology of Human Lumbar Plexus Roots Associated With
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
The Structure and Function of Breathing
CHAPTERCONTENTS The structure-function continuum 1 Multiple Influences: biomechanical, biochemical and psychological 1 The structure and Homeostasis and heterostasis 2 OBJECTIVE AND METHODS 4 function of breathing NORMAL BREATHING 5 Respiratory benefits 5 Leon Chaitow The upper airway 5 Dinah Bradley Thenose 5 The oropharynx 13 The larynx 13 Pathological states affecting the airways 13 Normal posture and other structural THE STRUCTURE-FUNCTION considerations 14 Further structural considerations 15 CONTINUUM Kapandji's model 16 Nowhere in the body is the axiom of structure Structural features of breathing 16 governing function more apparent than in its Lung volumes and capacities 19 relation to respiration. This is also a region in Fascla and resplrstory function 20 which prolonged modifications of function - Thoracic spine and ribs 21 Discs 22 such as the inappropriate breathing pattern dis- Structural features of the ribs 22 played during hyperventilation - inevitably intercostal musculature 23 induce structural changes, for example involving Structural features of the sternum 23 Posterior thorax 23 accessory breathing muscles as well as the tho- Palpation landmarks 23 racic articulations. Ultimately, the self-perpetuat- NEURAL REGULATION OF BREATHING 24 ing cycle of functional change creating structural Chemical control of breathing 25 modification leading to reinforced dysfunctional Voluntary control of breathing 25 tendencies can become complete, from The autonomic nervous system 26 whichever direction dysfunction arrives, for Sympathetic division 27 Parasympathetic division 27 example: structural adaptations can prevent NANC system 28 normal breathing function, and abnormal breath- THE MUSCLES OF RESPIRATION 30 ing function ensures continued structural adap- Additional soft tissue influences and tational stresses leading to decompensation. -

Vertebral Column and Thorax
Introduction to Human Osteology Chapter 4: Vertebral Column and Thorax Roberta Hall Kenneth Beals Holm Neumann Georg Neumann Gwyn Madden Revised in 1978, 1984, and 2008 The Vertebral Column and Thorax Sternum Manubrium – bone that is trapezoidal in shape, makes up the superior aspect of the sternum. Jugular notch – concave notches on either side of the superior aspect of the manubrium, for articulation with the clavicles. Corpus or body – flat, rectangular bone making up the major portion of the sternum. The lateral aspects contain the notches for the true ribs, called the costal notches. Xiphoid process – variably shaped bone found at the inferior aspect of the corpus. Process may fuse late in life to the corpus. Clavicle Sternal end – rounded end, articulates with manubrium. Acromial end – flat end, articulates with scapula. Conoid tuberosity – muscle attachment located on the inferior aspect of the shaft, pointing posteriorly. Ribs Scapulae Head Ventral surface Neck Dorsal surface Tubercle Spine Shaft Coracoid process Costal groove Acromion Glenoid fossa Axillary margin Medial angle Vertebral margin Manubrium. Left anterior aspect, right posterior aspect. Sternum and Xyphoid Process. Left anterior aspect, right posterior aspect. Clavicle. Left side. Top superior and bottom inferior. First Rib. Left superior and right inferior. Second Rib. Left inferior and right superior. Typical Rib. Left inferior and right superior. Eleventh Rib. Left posterior view and left superior view. Twelfth Rib. Top shows anterior view and bottom shows posterior view. Scapula. Left side. Top anterior and bottom posterior. Scapula. Top lateral and bottom superior. Clavicle Sternum Scapula Ribs Vertebrae Body - Development of the vertebrae can be used in aging of individuals. -

Part 1 the Thorax ECA1 7/18/06 6:30 PM Page 2 ECA1 7/18/06 6:30 PM Page 3
ECA1 7/18/06 6:30 PM Page 1 Part 1 The Thorax ECA1 7/18/06 6:30 PM Page 2 ECA1 7/18/06 6:30 PM Page 3 Surface anatomy and surface markings The experienced clinician spends much of his working life relating the surface anatomy of his patients to their deep structures (Fig. 1; see also Figs. 11 and 22). The following bony prominences can usually be palpated in the living subject (corresponding vertebral levels are given in brackets): •◊◊superior angle of the scapula (T2); •◊◊upper border of the manubrium sterni, the suprasternal notch (T2/3); •◊◊spine of the scapula (T3); •◊◊sternal angle (of Louis) — the transverse ridge at the manubrio-sternal junction (T4/5); •◊◊inferior angle of scapula (T8); •◊◊xiphisternal joint (T9); •◊◊lowest part of costal margin—10th rib (the subcostal line passes through L3). Note from Fig. 1 that the manubrium corresponds to the 3rd and 4th thoracic vertebrae and overlies the aortic arch, and that the sternum corre- sponds to the 5th to 8th vertebrae and neatly overlies the heart. Since the 1st and 12th ribs are difficult to feel, the ribs should be enu- merated from the 2nd costal cartilage, which articulates with the sternum at the angle of Louis. The spinous processes of all the thoracic vertebrae can be palpated in the midline posteriorly, but it should be remembered that the first spinous process that can be felt is that of C7 (the vertebra prominens). The position of the nipple varies considerably in the female, but in the male it usually lies in the 4th intercostal space about 4in (10cm) from the midline. -

Clinical Presentations of Lumbar Disc Degeneration and Lumbosacral Nerve Lesions
Hindawi International Journal of Rheumatology Volume 2020, Article ID 2919625, 13 pages https://doi.org/10.1155/2020/2919625 Review Article Clinical Presentations of Lumbar Disc Degeneration and Lumbosacral Nerve Lesions Worku Abie Liyew Biomedical Science Department, School of Medicine, Debre Markos University, Debre Markos, Ethiopia Correspondence should be addressed to Worku Abie Liyew; [email protected] Received 25 April 2020; Revised 26 June 2020; Accepted 13 July 2020; Published 29 August 2020 Academic Editor: Bruce M. Rothschild Copyright © 2020 Worku Abie Liyew. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Lumbar disc degeneration is defined as the wear and tear of lumbar intervertebral disc, and it is mainly occurring at L3-L4 and L4-S1 vertebrae. Lumbar disc degeneration may lead to disc bulging, osteophytes, loss of disc space, and compression and irritation of the adjacent nerve root. Clinical presentations associated with lumbar disc degeneration and lumbosacral nerve lesion are discogenic pain, radical pain, muscular weakness, and cutaneous. Discogenic pain is usually felt in the lumbar region, or sometimes, it may feel in the buttocks, down to the upper thighs, and it is typically presented with sudden forced flexion and/or rotational moment. Radical pain, muscular weakness, and sensory defects associated with lumbosacral nerve lesions are distributed on -

Bone Grafts and Implants in Spine Surgery
CHAPTER 39 BONE GRAFTS AND IMPLANTS IN SPINE SURGERY Ken Hsu James F. Zucherman Arthur H. White Recent advances in both fusion tech of scaffolds, bridges, spacers, fillers of J • niques and instrumentation have defects, and replacements of bone lost. markedly facilitated the treatment of Immobilization of multiple motion seg spinaldisorders. Yet asignificantnumber ments is frequently necessary in the spine; of patients exist who continue to have great demands arc made on bone grafts. pseudarthroses. Despite the surgical ad In the lumbosacral spine, body weight vances the essentials ofa successful spinal and muscular forcesimpart loads equal to fusion still appear to be the effective ap three or four times body weight. It is plication of sound bone grafting princi not surprising that the highest rate of ples. These principles, along with the bone graft failure is seen in the lumbo techniques, problems, and complications sacral spine. Hence, the following is a associated with bone grafting, arc re discussion of technical problems, bio- viewed in this chapter. mechanical and physiologic character The loss ofbone in the spine often pre istics of bone grafts, and implants. sents serious difficulties not seen in other areas. The most favorable replacement THE AUTOGRAFT would still be a bone graft that fills the Autograft, or bone graft transplanted defect and becomes incorporated into the from one site to another in the same indi spine. However, the availability of ap vidual, is considered to be the most bio propriate bone to replace the loss is a logically suitable. Its advantages include: significant problem. Alternatives to bone 1. -

Netter's Anatomy Flash Cards – Section 7 – List 4Th Edition
Netter's Anatomy Flash Cards – Section 7 – List 4th Edition https://www.memrise.com/course/1577594/ Section 7 Lower Limb (72 cards) Plate 7-1 Hip (Coxal) Bone: Lateral View 1.1 Posterior superior iliac spine 1.2 Posterior inferior iliac spine 1.3 Greater sciatic notch 1.4 Body of ilium 1.5 Body of ischium 1.6 Ischial tuberosity 1.7 Pubic tubercle 1.8 Acetabulum 1.9 Iliac crest Plate 7-2 Hip (Coxal) Bone: Medial View 2.1 Wing (ala) of ilium (iliac fossa) 2.2 Pecten pubis (pectineal line) 2.3 Ramus of ischium 2.4 Lesser sciatic notch 2.5 Ischial spine 2.6 Articular surface (for sacrum) 2.7 Iliac tuberosity Plate 7-3 Hip Joint: Lateral View 3.1 Lunate (articular) surface of acetabulum 3.2 Articular cartilage 3.3 Head of femur 3.4 Ligament of head of femur (cut) 3.5 Obturator membrane 3.6 Acetabular labrum (fibrocartilaginous) Plate 7-4 Hip Joint: Anterior and Posterior Views 4.1 Iliofemoral ligament (Y ligament of Bigelow) 4.2 Pubofemoral ligament 4.3 Iliofemoral ligament 4.4 Ischiofemoral ligament Plate 7-5 Femur 5.1 Greater trochanter 5.2 Shaft (body) 5.3 Lateral epicondyle 5.4 Lateral condyle 5.5 Medial condyle 5.6 Medial epicondyle 5.7 Adductor tubercle 5.8 Linea aspera (Medial lip; Lateral lip) 5.9 Lesser trochanter 5.10 Intertrochanteric crest 5.11 Neck 5.12 Head Plate 7-6 Tibia and Fibula 6.1 Lateral condyle 6.2 Apex, Head, and Neck of fibula 6.3 Fibula 6.4 Lateral malleolus 6.5 Medial malleolus 6.6 Tibia 6.7 Tibial tuberosity 6.8 Medial condyle 6.9 Superior articular surfaces (medial and lateral facets) 6.10 Malleolar fossa of lateral -

Chapter 21 Fractures of the Upper Thoracic Spine: Approaches and Surgical Management
Chapter 21 Fractures of the Upper Thoracic Spine: Approaches and Surgical Management Sean D Christie, M.D., John Song, M.D., and Richard G Fessler, M.D., Ph.D. INTRODUCTION Fractures occurring in the thoracic region account for approximately 17 to 23% of all traumatic spinal fractures (1), with 22% of traumatic spinal fractures occurring between T1 and T4 (16). More than half of these fractures result in neurological injury, and almost three-quarters of those impaired suffer from complete paralysis. Obtaining surgical access to the anterior vertebral elements of the upper thoracic vertebrae (T1–T6) presents a unique anatomic challenge. The thoracic cage, which narrows significantly as it approaches the thoracic inlet, has an intimate association between the vertebral column and the superior mediastinal structures. The supraclavicular, transmanubrial, transthoracic, and lateral parascapular extrapleural approaches each provide access to the anterior vertebral elements of the upper thoracic vertebrae. However, each of these approaches has distinct advantages and disadvantages and their use should be tailored to each individual patient’s situation. This chapter reviews these surgical approaches. Traditional posterior approaches are illustrated in Figure 21.1, but will not be discussed in depth here. ANATOMIC CONSIDERATIONS AND STABILITY The upper thoracic spine possesses unique anatomic and biomechanical properties. The anterior aspects of the vertebral bodies are smaller than the posterior aspects, which contribute to the physiological kyphosis present in this region of the spine. Furthermore, this orientation results in a ventrally positioned axis of rotation, predisposing this region to compression injuries. The combination and interaction of the vertebral bodies, ribs, and sternum increase the inherent biomechanical stability of this segment of the spine to 2 to 3 times that of the thoracolumbar junction. -

33. Spinal Nerves. Cervical Plexus
GUIDELINES Students’ independent work during preparation to practical lesson Academic discipline HUMAN ANATOMY Topic SPINAL NERVES. CERVICAL PLEXUS. 1. Relevance of the topic: The knowledge of structures of the peripheral nervous system, particularly cervical plexus and its branches is the base of clinical thinking in terms of differential diagnosis for the doctor of any specialty, but above all a neurologist, vertebroneurologist, traumatologist, dermatologist, general practitioner. 2. Specific objectives of practical lesson - Analyse the composition of fibres of anterior and posterior roots of spinal nerves. - Explain the formation of spinal nerve. - Suggest the definition of spinal nerve. - Classify spinal nerve branches. - Explain functional anatomy of thoracic spinal nerve branches. - Define term "plexus of somatic nerves" including the formation of cervical plexus. - Draw a scheme of spinal nerve: o а - in cervical region of spinal cord (except for the CVIII); o b - in thoracic region of spinal cord; o c - on the level of SII – SIV. - Analyse the connection of somatic nerve (thoracic spinal nerve) with ganglia of sympathetic trunk. - Create the conception of grey and white connecting branches in the functional aspect. 3. Basic level of preparation (interdisciplinary integration) of the student includes knowledge of medical biology and histology of the development of nervous system in phylogenesis and ontogenesis. Name of previous disciplines Obtained skills 1. Medical Biology and Histology Know ontogenesis and phylogenesis of nervous system. The structure of the neuron. 2. Sections of Human Anatomy: - osteology The student should have skills to describe the structure of - myology the spine in general, to be able to demonstrate structural features of the cervical vertebrae, their connections with each other and with the bones of the skull. -

The Blood Supply of the Lumbar and Sacral Plexuses in the Human Foetus* by M
J. Anat., Lond. (1964), 98, 1, 105-116 105 With 4 plates and 3 text-figures Printed in Great Britain The blood supply of the lumbar and sacral plexuses in the human foetus* BY M. H. DAYt Department of Anatomy, Royal Free Hospital School of Medicine INTRODUCTION The existence of a blood supply to peripheral nerve is well established. Recently, a number of authors have reviewed the literature of the field, among them Blunt (1956) and Abdullah (1958), who from their own observations have confirmed that peripheral nerves are supplied by regional vessels reinforcing longitudinally arranged channels which freely anastomose with each other. There is also evidence that posterior root ganglia are particularly well supplied with blood vessels (Abdullah, 1958), but the precise distribution and arrangement of arteries to some individual nerve trunks and plexuses is still in need of investigation. The literature reveals few references to the blood supply of the lumbar and sacral plexuses. The distribution of arteries to the roots and ganglia of the sacral nerves was noted by Haller (1756), but the most important contributions in this field were those of Bartholdy (1897) and Tonkoff (1898), whose observations on the lumbar and sacral plexuses form part of a general survey of the blood supply of peripheral nerve in man. They cited the lumbar, ilio-lumbar, median and lateral sacral arteries as well as the gluteal and pudendal vessels as sources of supply, but gave no indication of the frequency of these contributions. Subsequent authors including Hovelacque (1927), dealt briefly with the distribution of the lateral sacral, median sacral, gluteal and pudendal arteries to the sacral plexus, but treated more fully the blood supply of the sciatic nerve. -

The Supra-Iliac Anterior Quadratus Lumborum Block
Can J Anesth/J Can Anesth https://doi.org/10.1007/s12630-019-01312-z REPORTS OF ORIGINAL INVESTIGATIONS The supra-iliac anterior quadratus lumborum block: a cadaveric study and case series Le bloc du muscle carre´ des lombes ante´rieur par approche supra-iliaque : une e´tude cadave´rique et une se´rie de cas Hesham Elsharkawy, MD, MBA, MSc . Kariem El-Boghdadly, MBBS, BSc, FRCA, EDRA, MSc . Theresa J. Barnes, MD, MPH . Richard Drake, PhD . Kamal Maheshwari, MD, MPH . Loran Mounir Soliman, MD . Jean-Louis Horn, MD . Ki Jinn Chin, MD, FRCPC Received: 9 July 2018 / Revised: 10 December 2018 / Accepted: 10 December 2018 Ó Canadian Anesthesiologists’ Society 2019 Abstract Methods Ultrasound-guided bilateral supra-iliac anterior Purpose The local anesthetic injectate spread with fascial QL blocks were performed with 30 mL of India ink dye in plane blocks and corresponding clinical outcomes may six fresh adult cadavers. Cadavers were subsequently vary depending on the site of injection. We developed and dissected to determine distribution of the dye. In five evaluated a supra-iliac approach to the anterior quadratus patients undergoing hip surgery, a unilateral supra-iliac lumborum (QL) block and hypothesized that this single anterior QL block with 25 mL ropivacaine 0.5% followed injection might successfully block the lumbar and sacral by a continuous catheter infusion was performed. Patients plexus in cadavers and provide analgesia for patients were clinically assessed daily for block efficacy. undergoing hip surgery. Results The cadaveric injections showed consistent dye involvement of the majority of the branches of the lumbar plexus, including the femoral nerve, lateral femoral cutaneous nerve, ilioinguinal nerve, and iliohypogastric Permission to use images was obtained from the Cleveland Clinic nerve. -
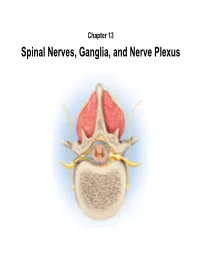
Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves
Chapter 13 Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves Posterior Spinous process of vertebra Posterior root Deep muscles of back Posterior ramus Spinal cord Transverse process of vertebra Posterior root ganglion Spinal nerve Anterior ramus Meningeal branch Communicating rami Anterior root Vertebral body Sympathetic ganglion Anterior General Anatomy of Nerves and Ganglia • Spinal cord communicates with the rest of the body by way of spinal nerves • nerve = a cordlike organ composed of numerous nerve fibers (axons) bound together by connective tissue – mixed nerves contain both afferent (sensory) and efferent (motor) fibers – composed of thousands of fibers carrying currents in opposite directions Anatomy of a Nerve Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Epineurium Perineurium Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Endoneurium Nerve Rootlets fiber Posterior root Fascicle Posterior root ganglion Anterior Blood root vessels Spinal nerve (b) Copyright by R.G. Kessel and R.H. Kardon, Tissues and Organs: A Text-Atlas of Scanning Electron Microscopy, 1979, W.H. Freeman, All rights reserved Blood vessels Fascicle Epineurium Perineurium Unmyelinated nerve fibers Myelinated nerve fibers (a) Endoneurium Myelin General Anatomy of Nerves and Ganglia • nerves of peripheral nervous system are ensheathed in Schwann cells – forms neurilemma and often a myelin sheath around the axon – external to neurilemma, each fiber is surrounded by -

Study of Anatomical Pattern of Lumbar Plexus in Human (Cadaveric Study)
54 Az. J. Pharm Sci. Vol. 54, September, 2016. STUDY OF ANATOMICAL PATTERN OF LUMBAR PLEXUS IN HUMAN (CADAVERIC STUDY) BY Prof. Gamal S Desouki, prof. Maged S Alansary,dr Ahmed K Elbana and Mohammad H Mandor FROM Professor Anatomy and Embryology Faculty of Medicine - Al-Azhar University professor of anesthesia Faculty of Medicine - Al-Azhar University Anatomy and Embryology Faculty of Medicine - Al-Azhar University Department of Anatomy and Embryology Faculty of Medicine of Al-Azhar University, Cairo Abstract The lumbar plexus is situated within the substance of the posterior part of psoas major muscle. It is formed by the ventral rami of the frist three nerves and greater part of the fourth lumbar nerve with or without a contribution from the ventral ramus of last thoracic nerve. The pattern of formation of lumbar plexus is altered if the plexus is prefixed (if the third lumbar is the lowest nerve which enters the lumbar plexus) or postfixed (if there is contribution from the 5th lumbar nerve). The branches of the lumbar plexus may be injured during lumbar plexus block and certain surgical procedures, particularly in the lower abdominal region (appendectomy, inguinal hernia repair, iliac crest bone graft harvesting and gynecologic procedures through transverse incisions). Thus, a better knowledge of the regional anatomy and its variations is essential for preventing the lesions of the branches of the lumbar plexus. Key Words: Anatomical variations, Lumbar plexus. Introduction The lumbar plexus formed by the ventral rami of the upper three nerves and most of the fourth lumbar nerve with or without a contribution from the ventral ramous of last thoracic nerve.