Current Status and Future Prospects of Perinatal Stem Cells
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Histopathological Analysis of Patients with Abruptio
HISTOPATHOLOGICAL ANALYSIS OF PATIENTS WITH ABRUPTIO PLACENTA IN A TERTIARY HEALTH CARE CENTRE Dissertation submitted in Partial fulfillment of the regulations required for the award of M.D. Degree in PATHOLOGY - BRANCH III THE TAMILNADU DR.M.G.R.MEDICAL UNIVERSITY CHENNAI MAY 2019 DECLARATION I hereby declare that the dissertation entitled “HISTOPATHOLOGICAL ANALYSIS OF PATIENTS WITH ABRUPTIO PLACENTA IN A TERTIARY HEALTH CARE CENTRE” is a bonafide research work done by me in the Department of Pathology, Coimbatore Medical College during the period from JANUARY 2017 TO JUNE 2018 under the guidance and supervision of Dr.G.S.THIRIVENI BALAJJI, M.D, Associate Professor, Department of Pathology, Coimbatore Medical College. This dissertation is submitted to The Tamilnadu Dr.MGR Medical University, Chennai towards the partial fulfillment of the requirement for the award of M.D., Degree (Branch III) in Pathology. I have not submitted this dissertation on any previous occasion to any University for the award of any Degree. Place : Coimbatore Dr. A. PETER SAMIDOSS Date : CERTIFICATE This is to certify that dissertation entitled “HISTOPATHOLOGICAL ANALYSIS OF PATIENTS WITH ABRUPTIO PLACENTA IN A TERTIARY HEALTH CARE CENTRE” is a bonafide work done by Dr. A. PETER SAMIDOSS, a postgraduate student in the Department of Pathology, Coimbatore Medical College, Coimbatore under guidance and supervision of Dr.G.S.THIRIVENI BALAJJI, M.D, Associate Professor, Department of Pathology, Coimbatore Medical College, Coimbatore in partial fulfillment of the regulations of the Tamil Nadu Dr. M. G. R. Medical University, Chennai towards the award of M.D. Degree (Branch III) in Pathology. Guide Head of the Department Dr.G.S.THIRIVENI BALAJJI, M.D, Prof. -

The Therapeutic Potential, Challenges and Future Clinical Directions of Stem Cells from the Wharton’S Jelly of the Human Umbilical Cord
Stem Cell Rev and Rep (2013) 9:226–240 DOI 10.1007/s12015-012-9418-z The Therapeutic Potential, Challenges and Future Clinical Directions of Stem Cells from the Wharton’s Jelly of the Human Umbilical Cord Ariff Bongso & Chui-Yee Fong Published online: 12 December 2012 # Springer Science+Business Media New York 2012 Abstract Mesenchymal stem cells (MSCs) from bone mar- attractive autologous or allogeneic agents for the treatment of row, adult organs and fetuses face the disadvantages of inva- malignant and non-malignant hematopoietic and non- sive isolation, limited cell numbers and ethical constraints hematopoietic diseases. This review critically evaluates their while embryonic stem cells (ESCs) and induced pluripotent therapeutic value, the challenges and future directions for their stem cells (iPSCs) face the clinical hurdles of potential immu- clinical application. norejection and tumorigenesis respectively. These challenges have prompted interest in the study and evaluation of stem Keywords Standardization of derivation protocols . cells from birth-associated tissues. The umbilical cord (UC) Properties and applications of Wharton’s jelly stem cells . has been the most popular. Hematopoietic stem cells (HSCs) Umbilical cord compartments harvested from cord blood have been successfully used for the treatment of hematopoietic diseases. Stem cell populations have also been reported in other compartments of the UC Introduction viz., amnion, subamnion, perivascular region, Wharton’sjelly, umbilical blood vessel adventia and endothelium. Differences Various types of stem cells have been isolated to date in the in stemness characteristics between compartments have been human from a variety of tissues including preimplantation reported and hence derivation protocols using whole UC embryos, fetuses, birth-associated tissues and adult organs. -

Messages from the Placentae Across Multiple Species a 50 Years
Placenta 84 (2019) 14–27 Contents lists available at ScienceDirect Placenta journal homepage: www.elsevier.com/locate/placenta Messages from the placentae across multiple species: A 50 years exploration T Hiroaki Soma Saitama Medical University, Japan ARTICLE INFO ABSTRACT Keywords: This review explores eight aspects of placentation in multiple mammalian. Gestational trophoblastic disease 1) Specialities of gestational trophoblastic disease. SUA(Single umbilical artery) 2) Clinical significance of single umbilical artery (SUA) syndrome. DIC(Disseminated intravascular coagulation) in 3) Pulmonary trophoblast embolism in pregnant chinchillas and DIC in pregnant giant panda. giant panda 4) Genetics status and placental behaviors during Japanese serow and related antelopes. Placentation in Japanese serow 5) Specific living style and placentation of the Sloth and Proboscis monkey. Hydatidiform mole in chimpanzee Placentation in different living elephant 6) Similarities of placental structures between human and great apes. Manatee and hyrax 7) Similarities of placental forms in elephants, manatees and rock hyrax with different living styles. Specific placental findings of Himalayan people 8) Specialities of placental pathology in Himalayan mountain people. Conclusions: It was taught that every mammalian species held on placental forms applied to different environ- mental life for their infants, even though their gestational lengths were different. 1. Introduction of effective chemotherapeutic agents. In 1959, I was fortunate tore- ceive an invitation from Prof. Kurt Benirschke at the Boston Lying-in Last October, Scientific American published a special issue about a Hospital. Before that, I had written to Prof. Arthur T. Hertig, Chairman baby's first organ, the placenta [1]. It is full of surprises and amazing of Pathology, Harvard Medical School, asking to study human tropho- science. -

FASD Effects of Alcohol on a Fetus
EFFECTS OF ALCOHOL ON A FETUS “Of all the substances of abuse (including cocaine, heroin, and marijuana), alcohol produces by far the most serious neurobehavioral effects in the fetus.” —Institute of Medicine Report to Congress, 19961 Prenatal exposure to alcohol can damage a fetus at any time, causing problems that persist throughout the individual’s life. There is no known safe level of alcohol use in pregnancy. WHAT IS THE SCOPE OF THE PROBLEM? identified in virtually every part of the body, including the brain, face, eyes, ears, heart, kidneys, and bones. No Alcohol is one of the most dangerous teratogens, which single mechanism can account for all the problems that are substances that can damage a developing fetus.1 Every alcohol causes. Rather, alcohol sets in motion many time a pregnant woman has a drink, her unborn child processes at different sites in the developing fetus: has one, too. Alcohol, like carbon monoxide from cigarettes, passes easily through the placenta from the • Alcohol can trigger cell death in a number of ways, mother's bloodstream into her baby's blood (See Figure causing different parts of the fetus to develop abnormally. 1)—and puts her fetus at risk of having a fetal alcohol • Alcohol can disrupt the way nerve cells develop, travel spectrum disorder (FASD). The blood alcohol level to form different parts of the brain, and function. (BAC) of the fetus becomes equal to or greater than the blood alcohol level of the mother. Because the fetus • By constricting the blood vessels, alcohol interferes with cannot break down alcohol the way an adult can, its BAC blood flow in the placenta, which hinders the delivery 2 remains high for a longer period of time. -
Glossary of Common MCH Terms and Acronyms
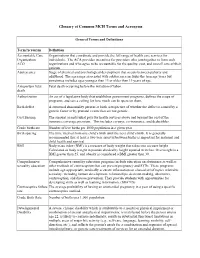
Glossary of Common MCH Terms and Acronyms General Terms and Definitions Term/Acronym Definition Accountable Care Organizations that coordinate and provide the full range of health care services for Organization individuals. The ACA provides incentives for providers who join together to form such ACO organizations and who agree to be accountable for the quality, cost, and overall care of their patients. Adolescence Stage of physical and psychological development that occurs between puberty and adulthood. The age range associated with adolescence includes the teen age years but sometimes includes ages younger than 13 or older than 19 years of age. Antepartum fetal Fetal death occurring before the initiation of labor. death Authorization An act of a legislative body that establishes government programs, defines the scope of programs, and sets a ceiling for how much can be spent on them. Birth defect A structural abnormality present at birth, irrespective of whether the defect is caused by a genetic factor or by prenatal events that are not genetic. Cost Sharing The amount an individual pays for health services above and beyond the cost of the insurance coverage premium. This includes co-pays, co-insurance, and deductibles. Crude birth rate Number of live births per 1000 population in a given year. Birth spacing The time interval from one child’s birth until the next child’s birth. It is generally recommended that at least a two-year interval between births is important for maternal and child health and survival. BMI Body mass index (BMI) is a measure of body weight that takes into account height. -

What Is the Role of the Placenta—Does It Protect Against Or Is It a Target for Insult?
Teratology Primer, 3rd Edition www.teratology.org/primer What Is the Role of the Placenta—Does It Protect Against or Is It a Target for Insult? Richard K. Miller University of Rochester School of Medicine & Dentistry Rochester, New York The placenta is not just a barrier but has many functions that are vital to the health of the embryo/fetus. The placenta is the anchor, the conduit, and the controller of pregnancy—and it can also be a target for toxicant action. The placenta encompasses not only the chorioallantoic placenta but all of its extraembryonic membranes (chorion/amnion) and the yolk sac. (Figure 1). The placenta and its membranes secure the embryo and fetus to the decidua (uterine lining) and release a variety of steroid and protein hormones that characterize the physiology of the pregnant female. Figure 1. Placental Structure in the Mouse. Figures depict early development of the mouse conceptus at embryonic days (E3.5, E7.5, E12.5). In the fetus, the visceral yolk sac (vYS) inverts and remains active throughout the entire gestation providing for transfer of selective large molecules, e.g., immunoglobulins IgG and vitamin B12. Abbreviations: Al, allantois; Am, amnion; Ch, chorion; Dec, decidua; Emb, embryo; Epc, ectoplacental cone; ICM, inner cell mass; Lab, Labyrinth; pYS, parietal yolk sac; SpT, spongiotrophoblast; TCG, trophoblast giant cell; Umb Cord, umbilical cord; vYS, visceral yolk sac; C-TGC, maternal blood canal trophoblast giant cell; P-TGC, parietal trophoblast giant cell; S-TGC, sinusoidal trophoblast giant cell; SpA-TGC, Spiral artery-associated trophoblast giant cell; Cyan-trophectoderm and trophoblast lineage, Black- inner cell mass and embryonic ectoderm; Gray -endoderm, Red-maternal vasculature, Purple-mesoderm, Yellow- decidua, Pink-fetal blood vessels in labyrinth. -

Equine Placenta – Marvelous Organ and a Lethal Weapon
Equine placenta – marvelous organ and a lethal weapon Malgorzata Pozor, DVM, PhD, Diplomate ACT Introduction Placenta has been defined as: „an apposition between parent (usually maternal) and fetal tissue in order to establish physiological exchange” (1). Another definition of this important organ was proposed by Steven and Morris: „a device consisting of one or more transport epithelia located between fetal and maternal blood supply” (2). The main function of placenta is to provide an interface between the dam and the the fetus and to allow the metabolic exchange of the the nutrients, oxygen and waste material. The maternal circulation is brought into a close apposition to the fetal circulation, while a separation of these two circulatory systems remain separated (3). A degree and complexity of this „intimate relationship” varies greately between species mostly due to the structural diversity of the extraembryonic membranes of the vertebrates. The early feto-maternal exchange in the equine pregnancy is established as early as on day 22 after fertilization. The fetal and choriovitellin circulations are already present, the capsule ruptures and the allantois is already visible (4). The allantois starts expanding by day 32 and vascularizes approximately 90% of the chorion and fuses with it to form chorioallantois by day 38 of gestation (5). The equine placenta continues increasing its complexity till approximately day 150 of gestation. Equids have epitheliochorial placenta, there are six leyers separating maternal and fetal circulation, and there are no erosion of the luminal, maternal epithelium, like in ruminants (6). Thousands of small chorionic microvilli develop and penetrate into endometrial invaginations. -

From Trophoblast to Human Placenta
From Trophoblast to Human Placenta (from The Encyclopedia of Reproduction) Harvey J. Kliman, M.D., Ph.D. Yale University School of Medicine I. Introduction II. Formation of the placenta III. Structure and function of the placenta IV. Complications of pregnancy related to trophoblasts and the placenta Glossary amnion the inner layer of the external membranes in direct contact with the amnionic fluid. chorion the outer layer of the external membranes composed of trophoblasts and extracellular matrix in direct contact with the uterus. chorionic plate the connective tissue that separates the amnionic fluid from the maternal blood on the fetal surface of the placenta. chorionic villous the final ramification of the fetal circulation within the placenta. cytotrophoblast a mononuclear cell which is the precursor cell of all other trophoblasts. decidua the transformed endometrium of pregnancy intervillous space the space in between the chorionic villi where the maternal blood circulates within the placenta invasive trophoblast the population of trophoblasts that leave the placenta, infiltrates the endo– and myometrium and penetrates the maternal spiral arteries, transforming them into low capacitance blood channels. Sunday, October 29, 2006 Page 1 of 19 From Trophoblasts to Human Placenta Harvey Kliman junctional trophoblast the specialized trophoblast that keep the placenta and external membranes attached to the uterus. spiral arteries the maternal arteries that travel through the myo– and endometrium which deliver blood to the placenta. syncytiotrophoblast the multinucleated trophoblast that forms the outer layer of the chorionic villi responsible for nutrient exchange and hormone production. I. Introduction The precursor cells of the human placenta—the trophoblasts—first appear four days after fertilization as the outer layer of cells of the blastocyst. -

Human Embryologyembryology
HUMANHUMAN EMBRYOLOGYEMBRYOLOGY Department of Histology and Embryology Jilin University ChapterChapter 22 GeneralGeneral EmbryologyEmbryology DevelopmentDevelopment inin FetalFetal PeriodPeriod 8.1 Characteristics of Fetal Period 210 days, from week 9 to delivery. characteristics: maturation of tissues and organs rapid growth of the body During 3-5 month, fetal growth in length is 5cm/M. In last 2 month, weight increases in 700g/M. relative slowdown in growth of the head compared with the rest of the body 8.2 Fetal AGE Fertilization age lasts 266 days, from the moment of fertilization to the day when the fetal is delivered. menstrual age last 280 days, from the first day of the last menstruation before pregnancy to the day when the fetal is delivered. The formula of expected date of delivery: year +1, month -3, day+7. ChapterChapter 22 GeneralGeneral EmbryologyEmbryology FetalFetal membranesmembranes andand placentaplacenta Villous chorion placenta Decidua basalis Umbilical cord Afterbirth/ secundines Fusion of amnion, smooth chorion, Fetal decidua capsularis, membrane decidua parietalis 9.1 Fetal Membranes TheThe fetalfetal membranemembrane includesincludes chorionchorion,, amnion,amnion, yolkyolk sac,sac, allantoisallantois andand umbilicalumbilical cord,cord, originatingoriginating fromfrom blastula.blastula. TheyThey havehave functionsfunctions ofof protection,protection, nutrition,nutrition, respiration,respiration, excretion,excretion, andand producingproducing hormonehormone toto maintainmaintain thethe pregnancy.pregnancy. delivery 1) Chorion: villous and smooth chorion Villus chorionic plate primary villus trophoblast secondary villus extraembryonic tertiary villus mesoderm stem villus Amnion free villus decidua parietalis Free/termin al villus Stem/ancho chorion ring villus Villous chorion Smooth chorion Amniotic cavity Extraembyonic cavity disappears gradually; Amnion is added into chorionic plate; Villous and smooth chorion is formed. -

Human Trophoblast-Endometrial Interactions in an in Vitro Suspension Culture System
Placenta (I99o), i I, 349-367 Human Trophoblast-Endometrial Interactions in an In Vitro Suspension Culture System HARVEY J. KLIMAN a,c, RONALD F. FEINBERG b & JULIA E. HAIMOWITZ a ~ Department of Pathology and Laboratory Medicine, University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania J9io4, USA b Department of Obstetrics and Gynecology, University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania 19io4, USA ' To whom all correspondence should be addressed at: Department of Pathology and Laboratory Medicine, 6 Founders Pavilion, University of Pennsylvania School of Medicine, 34oo Spruce Street, Philadelphia, Pennsylvania i9zo4-4283, USA Paper accepted 19.3.z99o SUMMARY We developed an in vitro suspension co-culture system to examine the interaction of ist, 2nd and 3rd trimester purified cytotrophoblasts with human endometrium. Endometrium explants were added to cytotrophoblast cell suspensions and placed on an angled gyrating platform in a 37~ incubator. When endometrium was cultured alone it was able to remain viable for up to 3 days. When trophoblasts were cultured alone, they formed small and large aggregates, and occasionally spherical shells with hollow centers. When trophoblasts and endometrium were cultured together, the trophoblasts adhered to the exposed stromal surfaces of the tissue frag- ments. The surface epithelium was not receptive to trophoblast attachment except in one ex- periment when day ~9 endometrium was used for the co-incubation, suggesting that surface attachment is usually restricted. A common finding was the presence of an acellular zone in the endometrium only adjacent to the attached trophoblasts. We speculate that this zone may be caused by proteolysis and resynthesis of ECM proteins by the trophoblasts. -

Abbreviations and Definitions
Appendix A Abbreviations and Definitions YYYY: 4 digit year FISH: Fluorescent In-situ Hybridization MM: 2 digit month FN2: FormsNet2 DD: 2 digit day FU: Follow-up AHOP: Adult, Hematology, Oncology or GVHD: Graft versus Host Disease Pediatric Unit (select only one) HSCT: Hematopoietic Stem Cell Transplant ALLO: Allogeneic HCT: Hematopoietic Cell Transplant ANC: Absolute Neutrophil Count KGF: Keratinocyte Growth Factor AUTO: Autologous NMDP: National Marrow Donor Program BM: Bone Marrow NOS: Not Otherwise Specified BMT-CTN: Blood & Marrow Transplant NST: Non-myeloablative Stem Cell Clinical Trials Network Transplant CIBMTR: Center for International Blood & PBSC: Peripheral Blood Stem Cells Marrow Transplant Research PCL: Plasma Cell Leukemia CIC: Center Identification Code PHI: Protected Health Information CMV: Cytomegalovirus Product Form: This was a transitional term CR: Complete Remission used to temporarily describe the form pieces CRF: Comprehensive Report Form = that came out of the ‘95/’02 CIBMTR Graft 2000 Baseline insert. The three Forms are IDM, HLA & 2004 IDM / 2005 HLA / 2006 INF INF. The term “Product Form” may have 20xx Disease (pre-HSCT) appeared in communication from Summer 2100 Day-100 2007; it is being retired. 2200 Six month- two year ProMISe: Electronic data collection system 2300 Greater than two year for EBMT 21xx Disease Follow-Up (Post-HSCT) PTLD: Posttransplant lymphoproliferative 2046 FNG / 2047, 2147 HEP / 2048,2148 disorder HIV RBC: Red Blood Cell 2900 Death RCI-BMT: Resource for Clinical CTN: Blood & Marrow -

Umbilical Cord Care N
n Umbilical Cord Care n (To keep your baby warm, baths should be quick during During pregnancy, the umbilical cord is the the first few weeks anyway.) baby’s connection to the mother. After birth, the cord is cut, and the stump remains connected to At every diaper change, use soap and water to clean the the baby. This is where the navel (belly button) will area around the cord stump. Pull up gently on the stump be. You’ll be taught some simple steps to care for and clean around its base. A little foul odor and moisture the cord stump after taking your baby home. are normal in this area. Although there are a few things to watch out for, Some doctors may recommend cleaning the stump with problems with the umbilical cord stump are rare. rubbing alcohol. You may clean the area with a small It usually falls off within a few weeks. amount of rubbing alcohol on a cotton ball or swab or use packaged alcohol wipes. Try not to use more rubbing alcohol than is necessary. Keep the cord stump over the diaper; it should not be What is the umbilical cord? inside the diaper. You may need to fold down the top While your baby was in the womb, he or she was edge of the diaper. connected to you by two important organs: the placenta and the umbilical cord. The umbilical cord is the fetus’s Are there any possible problems? “life support system” during pregnancy; it contains blood vessels that provide oxygen and nutrition.