A Mechanism for Diversity in Warning Signals: Conspicuousness Versus Toxicity in Poison Frogs
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

NC2006 (Fauna) Doc
NC2006 (fauna) Doc. 6 (English only/Únicamente en inglés/Seulement en anglais) CONVENTION ON INTERNATIONAL TRADE IN ENDANGERED SPECIES OF WILD FAUNA AND FLORA ____________ Nomenclature Committee Fauna Lima (Peru), 10 July 2006 AMPHIBIAN REFERENCE UPDATE 1. This document has been submitted by the zoologist of the Nomenclature Committee. 2. An extract from the current online reference "D.R. FROST, Amphibian Species of the World, Version 3.0 as of 2004" (see following pages) has been produced for the CITES listed species. Comparing it to the Taxonomic Checklist of CITES listed Amphibians provided by the former zoological Co-Chair of the Nomenclature Committee which was based on the FROST online-version 2.21 of 2002 the following nomenclature changes have been identified: Present Reference as adopted under Resolution Frost (2004): Amphibian Species of the World Conf. 12.11 (Rev. CoP13) Epipedobates femoralis Allobates femoralis Epipedobates zaparo Allobates zaparo not included Dendrobates amazonicus not included Dendrobates duellmani not included Dendrobates flavovittatus Dendrobates labialis Epipedobates labialis not included Dendrobates rubrocephalus included in Dendrobates imitator Dendrobates sirensis included in Dendrobates ventrimaculatus Dendrobates variabilis included in Epipedobates tricolor Epipedobates anthonyi not included Epipedobates pongoensis Minyobates spp. all species transferred to genus Dendrobates 3. The participants in the meeting are invited to comment on the reference, which will be proposed for adoption at the 14th meeting of the Conference of the Parties. NC2006 (fauna) Doc. 6 – p. 1 NC2006 (fauna) Doc. 6 – p. 2 Taxonomic Checklist of CITES listed Amphibians - Draft - Information extracted from FROST, D. R. (2004) "Amphibian Species of the World, an online Reference" V. -

Species Diversity and Conservation Status of Amphibians in Madre De Dios, Southern Peru
Herpetological Conservation and Biology 4(1):14-29 Submitted: 18 December 2007; Accepted: 4 August 2008 SPECIES DIVERSITY AND CONSERVATION STATUS OF AMPHIBIANS IN MADRE DE DIOS, SOUTHERN PERU 1,2 3 4,5 RUDOLF VON MAY , KAREN SIU-TING , JENNIFER M. JACOBS , MARGARITA MEDINA- 3 6 3,7 1 MÜLLER , GIUSEPPE GAGLIARDI , LILY O. RODRÍGUEZ , AND MAUREEN A. DONNELLY 1 Department of Biological Sciences, Florida International University, 11200 SW 8th Street, OE-167, Miami, Florida 33199, USA 2 Corresponding author, e-mail: [email protected] 3 Departamento de Herpetología, Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos, Avenida Arenales 1256, Lima 11, Perú 4 Department of Biology, San Francisco State University, 1600 Holloway Avenue, San Francisco, California 94132, USA 5 Department of Entomology, California Academy of Sciences, 55 Music Concourse Drive, San Francisco, California 94118, USA 6 Departamento de Herpetología, Museo de Zoología de la Universidad Nacional de la Amazonía Peruana, Pebas 5ta cuadra, Iquitos, Perú 7 Programa de Desarrollo Rural Sostenible, Cooperación Técnica Alemana – GTZ, Calle Diecisiete 355, Lima 27, Perú ABSTRACT.—This study focuses on amphibian species diversity in the lowland Amazonian rainforest of southern Peru, and on the importance of protected and non-protected areas for maintaining amphibian assemblages in this region. We compared species lists from nine sites in the Madre de Dios region, five of which are in nationally recognized protected areas and four are outside the country’s protected area system. Los Amigos, occurring outside the protected area system, is the most species-rich locality included in our comparison. -

Mimicry - Ecology - Oxford Bibliographies 12/13/12 7:29 PM
Mimicry - Ecology - Oxford Bibliographies 12/13/12 7:29 PM Mimicry David W. Kikuchi, David W. Pfennig Introduction Among nature’s most exquisite adaptations are examples in which natural selection has favored a species (the mimic) to resemble a second, often unrelated species (the model) because it confuses a third species (the receiver). For example, the individual members of a nontoxic species that happen to resemble a toxic species may dupe any predators by behaving as if they are also dangerous and should therefore be avoided. In this way, adaptive resemblances can evolve via natural selection. When this phenomenon—dubbed “mimicry”—was first outlined by Henry Walter Bates in the middle of the 19th century, its intuitive appeal was so great that Charles Darwin immediately seized upon it as one of the finest examples of evolution by means of natural selection. Even today, mimicry is often used as a prime example in textbooks and in the popular press as a superlative example of natural selection’s efficacy. Moreover, mimicry remains an active area of research, and studies of mimicry have helped illuminate such diverse topics as how novel, complex traits arise; how new species form; and how animals make complex decisions. General Overviews Since Henry Walter Bates first published his theories of mimicry in 1862 (see Bates 1862, cited under Historical Background), there have been periodic reviews of our knowledge in the subject area. Cott 1940 was mainly concerned with animal coloration. Subsequent reviews, such as Edmunds 1974 and Ruxton, et al. 2004, have focused on types of mimicry associated with defense from predators. -

Thermal Adaptation of Amphibians in Tropical Mountains
Thermal adaptation of amphibians in tropical mountains. Consequences of global warming Adaptaciones térmicas de anfibios en montañas tropicales: consecuencias del calentamiento global Adaptacions tèrmiques d'amfibis en muntanyes tropicals: conseqüències de l'escalfament global Pol Pintanel Costa ADVERTIMENT. La consulta d’aquesta tesi queda condicionada a l’acceptació de les següents condicions d'ús: La difusió d’aquesta tesi per mitjà del servei TDX (www.tdx.cat) i a través del Dipòsit Digital de la UB (diposit.ub.edu) ha estat autoritzada pels titulars dels drets de propietat intel·lectual únicament per a usos privats emmarcats en activitats d’investigació i docència. No s’autoritza la seva reproducció amb finalitats de lucre ni la seva difusió i posada a disposició des d’un lloc aliè al servei TDX ni al Dipòsit Digital de la UB. No s’autoritza la presentació del seu contingut en una finestra o marc aliè a TDX o al Dipòsit Digital de la UB (framing). Aquesta reserva de drets afecta tant al resum de presentació de la tesi com als seus continguts. En la utilització o cita de parts de la tesi és obligat indicar el nom de la persona autora. ADVERTENCIA. La consulta de esta tesis queda condicionada a la aceptación de las siguientes condiciones de uso: La difusión de esta tesis por medio del servicio TDR (www.tdx.cat) y a través del Repositorio Digital de la UB (diposit.ub.edu) ha sido autorizada por los titulares de los derechos de propiedad intelectual únicamente para usos privados enmarcados en actividades de investigación y docencia. -

N.Orntates PUBLISHED by the AMERICAN MUSEUM of NATURAL HISTORY CENTRAL PARK WEST at 79TH STREET, NEW YORK, N.Y
AMERICAN MUSEUM N.orntates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N.Y. 10024 Number 3068, 15 pp., 12 figures, 1 table June 11, 1993 A New Poison Frog from Manu INational Park, Southeastern Peru (Dendrobatidae, Epipedobates) LILY RODRIGUEZ' AND CHARLES W. MYERS2 ABSTRACT Epipedobates macero is a new species of den- ilar to a few other species occurring along the An- drobatid poison frog from lowland rain forest of dean front in eastern Peru, namely E. petersi and the Manu National Park, in the upper Madre de E. cainarachi, which differ in details ofcoloration, Dios drainage ofsoutheastern Peru. It is most sim- morphology, and vocalization. RESUMEN Epipedobates macero, especie nueva, es un den- del llano amaz6nico al pie de los Andes orientales drobatido venenoso de la selva pluvial baja del peruanos, a saber, E. petersi y E. cainarachi, las Parque Nacional del Manu, en el drenaje del Rio cuales difieren en detalles de coloracion, morfo- Alto Madre de Dios, al sudeste del Peru. Es similar logla, y vocalizacion. a otras dos especies que ocurren en los bosques ' Field Associate, Department of Herpetology and Ichthyology, American Museum of Natural History. Investi- gadora Asociada: Asociaci6n Peruana para la Conservaci6n de la Naturaleza (APECO), Parque Jos6 de Acosta 187, Lima 17, Perfi; and Museo de Historia Natural de la Universidad Mayor de San Marcos, apartado 140434, Lima 14, Peru. 2 Curator, Department of Herpetology and Ichthyology, American Museum of Natural History. Copyright © American Museum of Natural History 1993 ISSN 0003-0082 / Price $3.90 2 AMERICAN MUSEUM NOVITATES NO. -

Rapid Evolution of Mimicry Following Local Model Extinction Rsbl.Royalsocietypublishing.Org Christopher K
Evolutionary biology Rapid evolution of mimicry following local model extinction rsbl.royalsocietypublishing.org Christopher K. Akcali and David W. Pfennig Department of Biology, University of North Carolina, Chapel Hill, NC 27599-3280, USA Research Batesian mimicry evolves when individuals of a palatable species gain the selective advantage of reduced predation because they resemble a toxic Cite this article: Akcali CK, Pfennig DW. 2014 species that predators avoid. Here, we evaluated whether—and in which Rapid evolution of mimicry following local direction—Batesian mimicry has evolved in a natural population of model extinction. Biol. Lett. 10: 20140304. mimics following extirpation of their model. We specifically asked whether http://dx.doi.org/10.1098/rsbl.2014.0304 the precision of coral snake mimicry has evolved among kingsnakes from a region where coral snakes recently (1960) went locally extinct. We found that these kingsnakes have evolved more precise mimicry; by contrast, no such change occurred in a sympatric non-mimetic species or in conspecifics Received: 9 April 2014 from a region where coral snakes remain abundant. Presumably, more pre- cise mimicry has continued to evolve after model extirpation, because Accepted: 22 May 2014 relatively few predator generations have passed, and the fitness costs incurred by predators that mistook a deadly coral snake for a kingsnake were historically much greater than those incurred by predators that mistook a kingsnake for a coral snake. Indeed, these results are consistent with prior Subject Areas: theoretical and empirical studies, which revealed that only the most precise evolution, ecology, behaviour mimics are favoured as their model becomes increasingly rare. -

Volume 2. Animals
AC20 Doc. 8.5 Annex (English only/Seulement en anglais/Únicamente en inglés) REVIEW OF SIGNIFICANT TRADE ANALYSIS OF TRADE TRENDS WITH NOTES ON THE CONSERVATION STATUS OF SELECTED SPECIES Volume 2. Animals Prepared for the CITES Animals Committee, CITES Secretariat by the United Nations Environment Programme World Conservation Monitoring Centre JANUARY 2004 AC20 Doc. 8.5 – p. 3 Prepared and produced by: UNEP World Conservation Monitoring Centre, Cambridge, UK UNEP WORLD CONSERVATION MONITORING CENTRE (UNEP-WCMC) www.unep-wcmc.org The UNEP World Conservation Monitoring Centre is the biodiversity assessment and policy implementation arm of the United Nations Environment Programme, the world’s foremost intergovernmental environmental organisation. UNEP-WCMC aims to help decision-makers recognise the value of biodiversity to people everywhere, and to apply this knowledge to all that they do. The Centre’s challenge is to transform complex data into policy-relevant information, to build tools and systems for analysis and integration, and to support the needs of nations and the international community as they engage in joint programmes of action. UNEP-WCMC provides objective, scientifically rigorous products and services that include ecosystem assessments, support for implementation of environmental agreements, regional and global biodiversity information, research on threats and impacts, and development of future scenarios for the living world. Prepared for: The CITES Secretariat, Geneva A contribution to UNEP - The United Nations Environment Programme Printed by: UNEP World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge CB3 0DL, UK © Copyright: UNEP World Conservation Monitoring Centre/CITES Secretariat The contents of this report do not necessarily reflect the views or policies of UNEP or contributory organisations. -

*For More Information, Please See
Common Name Scientific Name Health Point Specifies-Specific Course(s)* Bat, Frog-eating Trachops cirrhosus AN0023 3198 3928 Bat, Fruit - Jamaican Artibeus jamaicensis AN0023 3198 3928 Bat, Mexican Free-tailed Tadarida brasiliensis mexicana AN0023 3198 3928 Bat, Round-eared - stripe-headed Tonatia saurophila AN0023 3198 3928 Bat, Round-eared - white-throated Lophostoma silvicolum AN0023 3198 3928 Bat, Seba's short-tailed Carollia perspicillata AN0023 3198 3928 Bat, Vampire - Common Desmodus rotundus AN0023 3198 3928 Bat, Vampire - Lesser False Megaderma spasma AN0023 3198 3928 Bird, Blackbird - Red-winged Agelaius phoeniceus AN0020 3198 3928 Bird, Brown-headed Cowbird Molothurus ater AN0020 3198 3928 Bird, Chicken Gallus gallus AN0020 3198 3529 Bird, Duck - Domestic Anas platyrhynchos AN0020 3198 3928 Bird, Finch - House Carpodacus mexicanus AN0020 3198 3928 Bird, Finch - Zebra Taeniopygia guttata AN0020 3198 3928 Bird, Goose - Domestic Anser anser AN0020 3198 3928 Bird, Owl - Barn Tyto alba AN0020 3198 3928 Bird, Owl - Eastern Screech Megascops asio AN0020 3198 3928 Bird, Pigeon Columba livia AN0020 3198 3928 Bird, Quail - Japanese Coturnix coturnix japonica AN0020 3198 3928 Bird, Sparrow - Harris' Zonotrichia querula AN0020 3198 3928 Bird, Sparrow - House Passer domesticus AN0020 3198 3928 Bird, Sparrow - White-crowned Zonotrichia leucophrys AN0020 3198 3928 Bird, Sparrow - White-throated Zonotrichia albicollis AN0020 3198 3928 Bird, Starling - Common Sturnus vulgaris AN0020 3198 3928 Cat Felis domesticus AN0020 3198 279 Cow Bos taurus -

Adaptations for Survival: Symbioses, Camouflage & Mimicry
Adaptations for Survival: Symbioses, Camouflage & Mimicry OCN 201 Biology Lecture 11 http://www.berkeley.edu/news/media/releases/2005/03/24_octopus.shtml Symbiosis • Parasitism - negative effect on host • Commensalism - no effect on host • Mutualism - both parties benefit Often involves food but benefits may also include protection from predators, dispersal, or habitat Parasitism Leeches (Segmented Worms) Tongue Louse (Crustacean) Nematodes (Roundworms) Commensalism or Mutualism? Anemone shrimp http://magma.nationalgeographic.com/ Anemone fish http://www.scuba-equipment-usa.com/marine/APR04/ Mutualism Cleaner Shrimp and Eel http://magma.nationalgeographic.com/ Whale Barnacles & Lice What kinds of symbioses are these? Commensal Parasite Camouflage • Often important for predators and prey to avoid being seen • Predators to catch their prey and prey to hide from their predators • Camouflage: Passive or adaptive Passive Camouflage Countershading Sharks Birds Countershading coloration of the Caribbean reef shark © George Ryschkewitsch Fish JONATHAN CHESTER Mammals shiftingbaselines.org/blog/big_tuna.jpg http://www.nmfs.noaa.gov/pr/images/cetaceans/orca_spyhopping-noaa.jpg Passive Camouflage http://www.cspangler.com/images/photos/aquarium/weedy-sea-dragon2.jpg Adaptive Camouflage Camouflage by Accessorizing Decorator crab Friday Harbor Marine Health Observatory http://www.projectnoah.org/ Camouflage by Mimicry http://www.berkeley.edu/news/media/releases/2005/03/24_octopus.shtml Mimicry • Animals can gain protection (or even access to prey) by looking -

Episodic Correlations in Behavioural Lateralization Differ Between a Poison Frog and Its Mimic
Animal Behaviour xxx (xxxx) xxx Contents lists available at ScienceDirect Animal Behaviour journal homepage: www.elsevier.com/locate/anbehav Episodic correlations in behavioural lateralization differ between a poison frog and its mimic * Hannah M. Anderson a, , David N. Fisher a, b, Brendan L. McEwen a, Justin Yeager c, Jonathan N. Pruitt a, James B. Barnett a a Department of Psychology, Neuroscience & Behaviour, McMaster University, Hamilton, ON, Canada b School of Biological Sciences, University of Aberdeen, Aberdeen, U.K. c Biodiversidad Medio Ambiente y Salud (BIOMAS), Universidad de Las Americas, Quito, Ecuador article info Sensory and behavioural lateralization is thought to increase neural efficiency and facilitate coordinated Article history: behaviour across much of the animal kingdom. Complementary laterality, when tasks are lateralized to Received 5 August 2020 opposite sides, can increase the efficiency of multitasking, but predictable behaviour may increase Initial acceptance 17 September 2020 predation risk. Laterality is, however, variable in its scale, existing at both the population and individual Final acceptance 23 November 2020 level. Population level lateralization is thought to facilitate coordination of social behaviours whereas Available online xxx individual level biases may promote behavioural efficiency. We studied behavioural lateralization in MS. number: A20-00601R sympatric wild populations of two terrestrial frog species: the Ecuador poison frog, Ameerega bilinguis, and its Batesian mimic the sanguine poison -
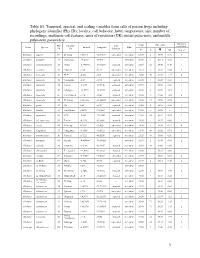
1 Table S1. Temporal, Spectral, and Scaling Variables from Calls Of
Table S1. Temporal, spectral, and scaling variables from calls of poison frogs including phylogeny identifier (Phy ID), locality, call behavior, habit, temperature, size, number of recordings, multinote call features, units of repetition (UR), initial pulse-note, and middle pulse-note parameters. Analyzed Phy Locality Call Temp SVL (mm) Genus Species Latitude Longitude Habit recordings ID ID Behavior °C N ! SD N of ♂ Allobates algorei 60 El Tama 7.65375 -72.19137 concealed terrestrial 23.50 8 18.90 0.70 3 Allobates brunneus 37 Guimaraes -15.2667 -55.5311 -- terrestrial 26.50 1 16.13 0.00 1 Allobates caeruleodactylus 48 Borba -4.398593 -59.60251 exposed terrestrial 25.60 12 15.50 0.40 1 Allobates crombiei 52 Altamira -3.65 -52.38 concealed terrestrial 24.10 2 18.10 0.04 2 Allobates femoralis 43 ECY -0.633 -76.5 concealed terrestrial 25.60 20 23.58 1.27 6 Allobates femoralis 46 Porongaba -8.67 -72.78 exposed terrestrial 25.00 1 25.38 0.00 1 Allobates femoralis 44 Leticia -4.2153 -69.9406 exposed terrestrial 25.50 1 20.90 0.00 1 Allobates femoralis 40 Albergue -12.8773 -71.3865 exposed terrestrial 26.00 6 21.98 2.18 1 Allobates femoralis 41 CAmazonico -12.6 -70.08 exposed terrestrial 26.00 12 22.43 1.06 4 Allobates femoralis 45 El Palmar 8.333333 -61.66667 concealed terrestrial 24.00 27 25.50 0.76 1 Allobates granti 49 FG 3.62 -53.17 exposed terrestrial 24.60 8 16.15 0.55 1 Allobates humilis 59 San Ramon 8.8678 -70.4861 concealed terrestrial 19.50 -- 21.80 -- 1 Allobates insperatus 54 ECY -0.633 -76.4005 exposed terrestrial 24.60 18 16.64 0.93 7 Allobates aff. -

Amphibia, Anura, Dendrobatidae, Allobates Femoralis (Boulenger, 1884): First Confirmed Country Records, Venezuela
ISSN 1809-127X (online edition) © 2010 Check List and Authors Chec List Open Access | Freely available at www.checklist.org.br Journal of species lists and distribution N Allobates femoralis ISTRIBUTIO Amphibia, Anura, Dendrobatidae, D Venezuela (Boulenger, 1884): First confirmed country records, RAPHIC G César L. Barrio-Amorós and Juan Carlos Santos 2 EO 1* G N O 1 Fundación Andígena, [email protected] postal 210, 5101-A. Mérida, Venezuela. OTES 2 University of Texas at Austin, Integrative Biology. 1 University Station C0930. Austin, TX 78705, USA. N * Corresponding author E-mail: Abstract: Allobates femoralis analized advertisement calls taken at two Venezuelan localities. The presence of the dendrobatid frog in Venezuela is herein confirmed though recorded and Allobates femoralis is a common dendrobatid frog widely distributed throughout the Amazon and Guianas in Colombia, Ecuador, Peru, Bolivia, Brazil, French Guiana, series of four notes at Las Claritas is 0.52 sec; the dominant Suriname and Guyana (Lötters et al. frequency is at 2954 Hz and the fundamental frequency is error, based on KU (Natural History 2007). Museum, The firstUniversity report of this species from Venezuela (Duellman 1997) was in was later recognized as Ameerega guayanensis (Barrio- Amorósof Kansas, 2004). Lawrence, Ameerega Kansas, guayanensisUSA) 167335. was This describedspecimen 2004).for populations Today however, of eastern this Venezuelataxon validity and is some questionable authors considered it as valid (Schulte 1999; Barrio-Amorós A.(J.C. (picta) Santos guayanensis unpublished because data). itSince is theno validnomenclatural available act was yet performed, we herein use the combinationAllobates femoralis was removed from the Venezuelan checklist name for those populations.