Gene Flow and Genetic Admixture Across a Secondary Contact Zone Between Two Divergent Lineages of the Eurasian Green Woodpecker Picusviridis J.-M
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Detecting and Managing Suspected Admixture and Genetic Drift in Domestic Livestock: Modern Dexter Cattle - a Case Study
Detecting and managing suspected admixture and genetic drift in domestic livestock: modern Dexter cattle - a case study Timothy C Bray Cardiff University C a r d if f UNIVERSITY PRIFYSCOL C a e RDY|§> A dissertation submitted to Cardiff University in candidature for the degree of Doctor of Philosophy UMI Number: U585124 All rights reserved INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed, a note will indicate the deletion. Dissertation Publishing UMI U585124 Published by ProQuest LLC 2013. Copyright in the Dissertation held by the Author. Microform Edition © ProQuest LLC. All rights reserved. This work is protected against unauthorized copying under Title 17, United States Code. ProQuest LLC 789 East Eisenhower Parkway P.O. Box 1346 Ann Arbor, Ml 48106-1346 Table of Contents Page Number Abstract I Declaration II Acknowledgements III Table of Contents IV Chapter 1. Introduction 1 1. introduction 2 1.1. Molecular genetics in conservation 2 1.2. Population genetic diversity 3 1.2.1. Microsatellites 3 1.2.2. Within-population variability 4 1.2.3. Population bottlenecks 5 1.2.4. Population differentiation 6 1.3. Assignment of conservation value 8 1.4. Genetic admixture 10 1.4.1. Admixture affecting conservation 12 1.5. Quantification of admixture 13 1.5.1. Different methods of determining admixture proportions 14 1.5.1.1. Gene identities 16 1.5.1.2. -

The Genetic Reinscription of Race Author(S): Nadia Abu El-Haj Reviewed Work(S): Source: Annual Review of Anthropology, Vol. 36 (2007), Pp
The Genetic Reinscription of Race Author(s): Nadia Abu El-Haj Reviewed work(s): Source: Annual Review of Anthropology, Vol. 36 (2007), pp. 283-300 Published by: Annual Reviews Stable URL: http://www.jstor.org/stable/25064957 Accessed: 04/03/2013 00:08 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at http://www.jstor.org/page/info/about/policies/terms.jsp JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. Annual Reviews is collaborating with JSTOR to digitize, preserve and extend access to Annual Review of Anthropology. STOR http://www.jstor.org This content downloaded on Mon, 4 Mar 2013 00:08:07 AM All use subject to JSTOR Terms and Conditions The Genetic Reinscription of Race Nadia Abu El-Haj Department of Anthropology, Barnard College, Columbia University, New York, NY 10027: email: ne2(X)[email protected] Annu. Rev. Anthropol. 2007. 36:283-300 Key Words The. tnnii.il Rnritu tfAatbnfibgf is online at genomics, postgenomics, neo-liberalism, identity politics, risk, anthro.anruulrevtews.org biological citizenship I his ankle's tiul: Abstract 10.114rVannurev.anthm.34.081 8(14.120522 (Copyright © 2007 by Annual Reviews Critics have debated for the past decade or more whether race is All rights reserved dead or alive in "the new genetics": Is genomics opening up novel OOX4-6s-(i/()7/1021-0283S20.00 terrains for social identities or is it reauthorizing race? I explore the relationship between race and the new genetics by considering whether this "race" is the same scientific object as that produced by race science and whether these race-making practices are animated In similar social and political logics. -

European Red List of Birds
European Red List of Birds Compiled by BirdLife International Published by the European Commission. opinion whatsoever on the part of the European Commission or BirdLife International concerning the legal status of any country, Citation: Publications of the European Communities. Design and layout by: Imre Sebestyén jr. / UNITgraphics.com Printed by: Pannónia Nyomda Picture credits on cover page: Fratercula arctica to continue into the future. © Ondrej Pelánek All photographs used in this publication remain the property of the original copyright holder (see individual captions for details). Photographs should not be reproduced or used in other contexts without written permission from the copyright holder. Available from: to your questions about the European Union Freephone number (*): 00 800 6 7 8 9 10 11 (*) Certain mobile telephone operators do not allow access to 00 800 numbers or these calls may be billed Published by the European Commission. A great deal of additional information on the European Union is available on the Internet. It can be accessed through the Europa server (http://europa.eu). Cataloguing data can be found at the end of this publication. ISBN: 978-92-79-47450-7 DOI: 10.2779/975810 © European Union, 2015 Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale or other commercial purposes is prohibited without prior written permission of the copyright holder. Printed in Hungary. European Red List of Birds Consortium iii Table of contents Acknowledgements ...................................................................................................................................................1 Executive summary ...................................................................................................................................................5 1. -

Phylogeography of the Eurasian Green Woodpecker (Picus Viridis)
Journal of Biogeography (J. Biogeogr.) (2011) 38, 311–325 ORIGINAL Phylogeography of the Eurasian green ARTICLE woodpecker (Picus viridis) J.-M. Pons1,2*, G. Olioso3, C. Cruaud4 and J. Fuchs5,6 1UMR7205 ‘Origine, Structure et Evolution de ABSTRACT la Biodiversite´’, De´partement Syste´matique et Aim In this paper we investigate the evolutionary history of the Eurasian green Evolution, Muse´um National d’Histoire Naturelle, 55 rue Buffon, C.P. 51, 75005 Paris, woodpecker (Picus viridis) using molecular markers. We specifically focus on the France, 2Service Commun de Syste´matique respective roles of Pleistocene climatic oscillations and geographical barriers in Mole´culaire, IFR CNRS 101, Muse´um shaping the current population genetics within this species. In addition, we National d’Histoire Naturelle, 43 rue Cuvier, discuss the validity of current species and subspecies limits. 3 75005 Paris, France, 190 rue de l’industrie, Location Western Palaearctic: Europe to western Russia, and Africa north of the 4 11210 Port la Nouvelle, France, Genoscope, Sahara. Centre National de Se´quenc¸age, 2 rue Gaston Cre´mieux, CP5706, 91057 Evry Cedex, France, Methods We sequenced two mitochondrial genes and five nuclear introns for 17 5DST/NRF Centre of Excellence at the Percy Eurasian green woodpeckers. Multilocus phylogenetic analyses were conducted FitzPatrick Institute, University of Cape Town, using maximum likelihood and Bayesian algorithms. In addition, we sequenced a Rondebosch 7701, Cape Town, South Africa, fragment of the cytochrome b gene (cyt b, 427 bp) and of the Z-linked BRM 6Museum of Vertebrate Zoology and intron 15 for 113 and 85 individuals, respectively. The latter data set was analysed Department of Integrative Biology, 3101 Valley using population genetic methods. -
ORL 5.1 Non-Passerines Final Draft01a.Xlsx
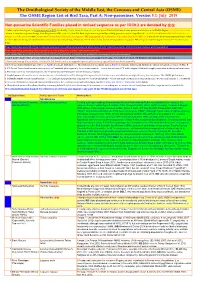
The Ornithological Society of the Middle East, the Caucasus and Central Asia (OSME) The OSME Region List of Bird Taxa, Part A: Non-passerines. Version 5.1: July 2019 Non-passerine Scientific Families placed in revised sequence as per IOC9.2 are denoted by ֍֍ A fuller explanation is given in Explanation of the ORL, but briefly, Bright green shading of a row (eg Syrian Ostrich) indicates former presence of a taxon in the OSME Region. Light gold shading in column A indicates sequence change from the previous ORL issue. For taxa that have unproven and probably unlikely presence, see the Hypothetical List. Red font indicates added information since the previous ORL version or the Conservation Threat Status (Critically Endangered = CE, Endangered = E, Vulnerable = V and Data Deficient = DD only). Not all synonyms have been examined. Serial numbers (SN) are merely an administrative convenience and may change. Please do not cite them in any formal correspondence or papers. NB: Compass cardinals (eg N = north, SE = southeast) are used. Rows shaded thus and with yellow text denote summaries of problem taxon groups in which some closely-related taxa may be of indeterminate status or are being studied. Rows shaded thus and with yellow text indicate recent or data-driven major conservation concerns. Rows shaded thus and with white text contain additional explanatory information on problem taxon groups as and when necessary. English names shaded thus are taxa on BirdLife Tracking Database, http://seabirdtracking.org/mapper/index.php. Nos tracked are small. NB BirdLife still lump many seabird taxa. A broad dark orange line, as below, indicates the last taxon in a new or suggested species split, or where sspp are best considered separately. -

EUROPEAN BIRDS of CONSERVATION CONCERN Populations, Trends and National Responsibilities
EUROPEAN BIRDS OF CONSERVATION CONCERN Populations, trends and national responsibilities COMPILED BY ANNA STANEVA AND IAN BURFIELD WITH SPONSORSHIP FROM CONTENTS Introduction 4 86 ITALY References 9 89 KOSOVO ALBANIA 10 92 LATVIA ANDORRA 14 95 LIECHTENSTEIN ARMENIA 16 97 LITHUANIA AUSTRIA 19 100 LUXEMBOURG AZERBAIJAN 22 102 MACEDONIA BELARUS 26 105 MALTA BELGIUM 29 107 MOLDOVA BOSNIA AND HERZEGOVINA 32 110 MONTENEGRO BULGARIA 35 113 NETHERLANDS CROATIA 39 116 NORWAY CYPRUS 42 119 POLAND CZECH REPUBLIC 45 122 PORTUGAL DENMARK 48 125 ROMANIA ESTONIA 51 128 RUSSIA BirdLife Europe and Central Asia is a partnership of 48 national conservation organisations and a leader in bird conservation. Our unique local to global FAROE ISLANDS DENMARK 54 132 SERBIA approach enables us to deliver high impact and long term conservation for the beneit of nature and people. BirdLife Europe and Central Asia is one of FINLAND 56 135 SLOVAKIA the six regional secretariats that compose BirdLife International. Based in Brus- sels, it supports the European and Central Asian Partnership and is present FRANCE 60 138 SLOVENIA in 47 countries including all EU Member States. With more than 4,100 staf in Europe, two million members and tens of thousands of skilled volunteers, GEORGIA 64 141 SPAIN BirdLife Europe and Central Asia, together with its national partners, owns or manages more than 6,000 nature sites totaling 320,000 hectares. GERMANY 67 145 SWEDEN GIBRALTAR UNITED KINGDOM 71 148 SWITZERLAND GREECE 72 151 TURKEY GREENLAND DENMARK 76 155 UKRAINE HUNGARY 78 159 UNITED KINGDOM ICELAND 81 162 European population sizes and trends STICHTING BIRDLIFE EUROPE GRATEFULLY ACKNOWLEDGES FINANCIAL SUPPORT FROM THE EUROPEAN COMMISSION. -

Models, Methods and Tools for Ancestry Inference and Admixture Analysis
Quantitative Biology 2017, 5(3): 236–250 DOI 10.1007/s40484-017-0117-2 REVIEW Models, methods and tools for ancestry inference and admixture analysis ,† ,† ,† Kai Yuan1,2 , Ying Zhou1,2 , Xumin Ni3 , Yuchen Wang1,2, Chang Liu1,2 and Shuhua Xu1,2,4,5,* 1 CAS Key Laboratory of Computational Biology, Max Planck Independent Research Group on Population Genomics, CAS-MPG Partner Institute for Computational Biology, Shanghai Institutes for Biological Sciences, CAS, Shanghai 200031, China 2 University of Chinese Academy of Sciences, Beijing 100049, China 3 Department of Mathematics, School of Science, Beijing Jiaotong University, Beijing 100044, China 4 School of Life Science and Technology, ShanghaiTech University, Shanghai 201210, China 5 Collaborative Innovation Center of Genetics and Development, Shanghai 200438, China * Correspondence: [email protected] Received May 5, 2017; Revised July 1, 2017; Accepted July 3, 2017 Background: Genetic admixture refers to the process or consequence of interbreeding between two or more previously isolated populations within a species. Compared to many other evolutionary driving forces such as mutations, genetic drift, and natural selection, genetic admixture is a quick mechanism for shaping population genomic diversity. In particular, admixture results in “recombination” of genetic variants that have been fixed in different populations, which has many evolutionary and medical implications. Results: However, it is challenging to accurately reconstruct population admixture history and to understand of population admixture dynamics. In this review, we provide an overview of models, methods, and tools for ancestry inference and admixture analysis. Conclusions: Many methods and tools used for admixture analysis were originally developed to analyze human data, but these methods can also be directly applied and/or slightly modified to study non-human species as well. -

Birding I 2016
Spain Trip Report 22nd April to 7th May 2016 Wallcreeper by Ray Wilson Trip report by tour leader: Mark Beevers and Alberto Bueno Top ten birds as voted for by participants: 1. Spanish Imperial Eagle 6. Great Bustard 2. Wallcreeper 7. Dupont’s Lark 3. Dotterel 8. Eagle Owl 4. Bearded Reedling 9. Tawny Owl 5. Little Bustard 10. Eurasian Penduline Tit RBT Trip Report Tour Name & Number 20YY 2 Tour Summary Day one and we set off on time from our rendezvous at the airport. It took a while to negotiate the Madrid traffic but by mid-morning we were heading south-west on remarkably quiet roads towards Monfragüe National Park with a couple of scheduled stops planned. Our first stop however was unscheduled, when Alberto saw a Hawfinch from the vehicle at Colmenar del Arroyo. The Hawfinch could not be relocated but this stop gave us an introduction to our first Mediterranean species with European Bee-eaters, Woodchat Shrike and Black- Redstarts all being seen along with a pair of Rock Sparrows. Our first Eurasian Griffon and Cinereous Bluethroat by Ignacio Yúfera Vultures were overhead and a pair of Short-toed Treecreepers showed very well. Not a bad first stop. Our first scheduled stop was at Navahonda Hermitage where after quite a bit of searching we eventually found a Hawfinch, which was our target here. We also secured great looks at a Nightingale, a species which we were to hear singing frequently during the tour. Eurasian Nuthatches were particularly common here, giving great views, whilst further Mediterranean species included Red- rumped Swallow and European Serin. -

A Genome-Wide Perspective on the Evolutionary History of Enigmatic Wolf-Like Canids
Downloaded from genome.cshlp.org on October 1, 2021 - Published by Cold Spring Harbor Laboratory Press Research A genome-wide perspective on the evolutionary history of enigmatic wolf-like canids Bridgett M. vonHoldt,1 John P. Pollinger,1 Dent A. Earl,2 James C. Knowles,1 Adam R. Boyko,3 Heidi Parker,4 Eli Geffen,5 Malgorzata Pilot,6 Wlodzimierz Jedrzejewski,7 Bogumila Jedrzejewska,7 Vadim Sidorovich,7 Claudia Greco,8 Ettore Randi,8 Marco Musiani,9 Roland Kays,10 Carlos D. Bustamante,3 Elaine A. Ostrander,4 John Novembre,1 and Robert K. Wayne1,11 1Department of Ecology and Evolutionary Biology, University of California, Los Angeles, California 90095, USA; 2Department of Biomolecular Engineering, University of California, Santa Cruz, California 95064, USA; 3Department of Genetics, Stanford School of Medicine, Stanford, California 94305, USA; 4Cancer Genetics Branch, National Human Genome Research Institute, National Institutes of Health, Bethesda, Maryland 20892, USA; 5Department of Zoology, Tel Aviv University, Tel Aviv 69978, Israel; 6Museum and Institute of Zoology, Polish Academy of Sciences, 00-679 Warszawa, Poland; 7Mammal Research Institute, Polish Academy of Sciences, 17-230 Bialowieza, Poland; 8Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA), 40064 Ozzano Emilia (BO), Italy; 9Faculty of Environmental Design, University of Calgary, Calgary, Alberta T2N 1N4, Canada; 10New York State Museum, CEC 3140, Albany, New York 12230, USA High-throughput genotyping technologies developed for model species can potentially increase the resolution of de- mographic history and ancestry in wild relatives. We use a SNP genotyping microarray developed for the domestic dog to assay variation in over 48K loci in wolf-like species worldwide. -

Spain 2019 Species List
Spain 2019 Leader: Javi Elorriaga and Eagle-Eye Tours Pablo Perez Martinez Species List BIRD SPECIES - 162 total Seen/ No. Common Name Latin Name Heard ANSERIFORMES: Anatidae 1 Graylag Goose Anser anser s 2 Mute Swan Cygnus olor s 3 Common Shelduck Tadorna tadorna s 4 Gadwall Anas strepera s 5 Northern Shoveler Anas clypeata s 6 Mallard Anas platyrhynchos s 7 Marbled Teal Marmaronetta angustirostris s 8 Red-crested Pochard Netta rufina s 9 Common Pochard Aythya ferina s 10 Ferruginous Duck Aythya nyroca s 11 White-headed Duck Oxyura leucocephala s GALLIFORMES: Phasianidae 12 Common Quail Coturnix coturnix s 13 Red-legged Partridge Alectoris rufa s PHOENICOPTERIFORMES: Phoenicopteridae 14 Greater Flamingo Phoenicopterus roseus s PODICIPEDIFORMES: Podicipedidae 15 Little GreBe Tachybaptus ruficollis s 16 Great Crested GreBe Podiceps cristatus s 17 Eared GreBe Podiceps nigricollis s COLUMBIFORMES: Columbidae 18 Rock Pigeon (feral type) Columba livia s 19 Common Wood-Pigeon Columba palumbus s 20 European Turtle-Dove Streptopelia turtur s 21 Eurasian Collared-Dove Streptopelia decaocto s PTEROCLIFORMES: Pteroclidae 22 Pin-tailed Sandgrouse Pterocles alchata s OTIDIFORMES: Otididae 23 Great Bustard Otis tarda s 24 Little Bustard Tetrax tetrax s CUCULIFORMES: Cuculidae 25 Great Spotted Cuckoo Clamator glandarius s 26 Common Cuckoo Cuculus canorus s CAPRIMULGIFORMES: Caprimulgidae 27 Red-necked Nightjar Caprimulgus ruficollis s CAPRIMULGIFORMES: Apodidae 28 Alpine Swift Tachymarptis melba s Page 1 of 6 Spain 2019 Leader: Javi Elorriaga and Eagle-Eye -

Insight Into the Genetic Population Structure of Wild Red Foxes in Poland Reveals Low Risk of Genetic Introgression from Escaped Farm Red Foxes
G C A T T A C G G C A T genes Article Insight into the Genetic Population Structure of Wild Red Foxes in Poland Reveals Low Risk of Genetic Introgression from Escaped Farm Red Foxes Heliodor Wierzbicki * , Magdalena Zato ´n-Dobrowolska , Anna Mucha and Magdalena Moska Department of Genetics, Wrocław University of Environmental and Life Sciences, 51-631 Wrocław, Poland; [email protected] (M.Z.-D.); [email protected] (A.M.); [email protected] (M.M.) * Correspondence: [email protected]; Tel.: +48-71-3205-780 Abstract: In this study we assessed the level of genetic introgression between red foxes bred on fur farms in Poland and the native wild population. We also evaluated the impact of a geographic barrier and isolation by distance on gene flow between two isolated subpopulations of the native red fox and their genetic differentiation. Nuclear and mitochondrial DNA was collected from a total of 308 individuals (200 farm and 108 wild red foxes) to study non-native allele flow from farm into wild red fox populations. Genetic structure analyses performed using 24 autosomal microsatellites showed two genetic clusters as being the most probable number of distinct populations. No strong admixture signals between farm and wild red foxes were detected, and significant genetic differentiation was Citation: Wierzbicki, H.; identified between the two groups. This was also apparent from the mtDNA analysis. None of the Zato´n-Dobrowolska, M.; Mucha, A.; concatenated haplotypes detected in farm foxes was found in wild animals. The consequence of Moska, M. -

E9.3. Updated ISAP for the European Roller
INTERNATIONAL SPECIES ACTION PLAN FOR THE EUROPEAN ROLLER CORACIAS GARRULUS GARRULUS revised version 2020 Prepared by: MME BirdLife Hungary on behalf of the European Commission Compilers Orsolya Kiss University of Szeged Timothée Schwartz A Rocha France Sanja Barisic Croatian Academy of Sciences and Arts, Institute of Ornithology Francisco Valera Institute of Ornithology Casa Béla Tokody MME BirdLife Hungary List of Contributors Albania Erald Xeka [email protected] University Of Tirana Austria Bernard Wieser [email protected] Lebende Erde im Vulkanland Michael Tiffenbach [email protected] Nature conservation Office- Styria Carina Nebel [email protected] Museum of Natural-History Vienna Belarus Maxim Tarantovich [email protected] Ivan Russkikh [email protected] Bulgaria Petar Iankov [email protected] BirdLife Bulgaria (BSPB) Croatia Sanja Barisic [email protected] Institute of Ornithology CASA Vesna Tutiš [email protected] Institute of Ornithology CASA Cyprus Christina Leronymidou [email protected] BirdLIfe Cyprus Czech Republic Petr Pavelcik [email protected] Lukas Viktora [email protected] Czech Society for Ornithology (CSO) England Tom Finch [email protected] University of East Anglia Ian Fisher [email protected] RSPB Simon Butler [email protected] University of East Anglia Phil Saunders [email protected] University of East Anglia France Timothée Schwartz [email protected] A Rocha France Germany Dr Tilman C. Schneider [email protected]