Tactile Stimulation on Adulthood Modifies the HPA Axis, Neurotrophic Factors, and GFAP Signaling Reverting Depression-Like Behavior in Female Rats
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
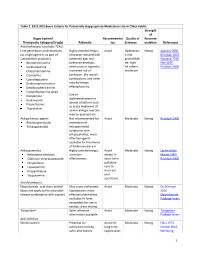
Table 2. 2012 AGS Beers Criteria for Potentially
Table 2. 2012 AGS Beers Criteria for Potentially Inappropriate Medication Use in Older Adults Strength of Organ System/ Recommendat Quality of Recomm Therapeutic Category/Drug(s) Rationale ion Evidence endation References Anticholinergics (excludes TCAs) First-generation antihistamines Highly anticholinergic; Avoid Hydroxyzin Strong Agostini 2001 (as single agent or as part of clearance reduced with e and Boustani 2007 combination products) advanced age, and promethazi Guaiana 2010 Brompheniramine tolerance develops ne: high; Han 2001 Carbinoxamine when used as hypnotic; All others: Rudolph 2008 Chlorpheniramine increased risk of moderate Clemastine confusion, dry mouth, Cyproheptadine constipation, and other Dexbrompheniramine anticholinergic Dexchlorpheniramine effects/toxicity. Diphenhydramine (oral) Doxylamine Use of diphenhydramine in Hydroxyzine special situations such Promethazine as acute treatment of Triprolidine severe allergic reaction may be appropriate. Antiparkinson agents Not recommended for Avoid Moderate Strong Rudolph 2008 Benztropine (oral) prevention of Trihexyphenidyl extrapyramidal symptoms with antipsychotics; more effective agents available for treatment of Parkinson disease. Antispasmodics Highly anticholinergic, Avoid Moderate Strong Lechevallier- Belladonna alkaloids uncertain except in Michel 2005 Clidinium-chlordiazepoxide effectiveness. short-term Rudolph 2008 Dicyclomine palliative Hyoscyamine care to Propantheline decrease Scopolamine oral secretions. Antithrombotics Dipyridamole, oral short-acting* May -

Management of Major Depressive Disorder Clinical Practice Guidelines May 2014
Federal Bureau of Prisons Management of Major Depressive Disorder Clinical Practice Guidelines May 2014 Table of Contents 1. Purpose ............................................................................................................................................. 1 2. Introduction ...................................................................................................................................... 1 Natural History ................................................................................................................................. 2 Special Considerations ...................................................................................................................... 2 3. Screening ........................................................................................................................................... 3 Screening Questions .......................................................................................................................... 3 Further Screening Methods................................................................................................................ 4 4. Diagnosis ........................................................................................................................................... 4 Depression: Three Levels of Severity ............................................................................................... 4 Clinical Interview and Documentation of Risk Assessment............................................................... -

Determination of Salivary Efavirenz by Liquid Chromatography Coupled with Tandem Mass Spectrometry
G Model CHROMB-17040; No. of Pages 5 ARTICLE IN PRESS Journal of Chromatography B, xxx (2010) xxx–xxx Contents lists available at ScienceDirect Journal of Chromatography B journal homepage: www.elsevier.com/locate/chromb Determination of salivary efavirenz by liquid chromatography coupled with tandem mass spectrometry Anri Theron a, Duncan Cromarty b, Malie Rheeders a, Michelle Viljoen a,∗ a Pharmacology, School of Pharmacy & Unit for Drug Research and Development North-West University, Private Bag X6001, Building G16, Room 113, Potchefstroom 2520, South Africa b Department of Pharmacology, Medical School, University of Pretoria, PO Box 2034, Pretoria 0001, South Africa article info abstract Article history: A novel and robust screening method for the determination of the non-nucleoside reverse transcriptase Received 12 May 2010 inhibitor, efavirenz (EFV), in human saliva has been developed and validated based on high perfor- Accepted 31 August 2010 mance liquid chromatography tandem mass spectrometry (LC–MS/MS). Sample preparation of the saliva Available online xxx involved solid-phase extraction (SPE) on C18 cartridges. The analytes were separated by high perfor- mance liquid chromatography (Phenomenex Kinetex C18, 150 mm × 3 mm internal diameter, 2.6 m Keywords: particle size) and detected with tandem mass spectrometry in electrospray positive ionization mode Efavirenz with multiple reaction monitoring. Gradient elution with increasing methanol (MeOH) concentration Saliva LC–MS/MS was used to elute the analytes, at a flow-rate of 0.4 mL/min. The total run time was 8.4 min and the retention times for the internal standard (reserpine) was 5.4 min and for EFV was 6.5 min. -

Phase I Safety, Pharmacokinetics, and Pharmacogenetics Study of the Antituberculosis Drug PA-824 with Concomitant Lopinavir-Ritonavir, Efavirenz, Or Rifampin
Phase I Safety, Pharmacokinetics, and Pharmacogenetics Study of the Antituberculosis Drug PA-824 with Concomitant Lopinavir-Ritonavir, Efavirenz, or Rifampin Kelly E. Dooley,a Anne F. Luetkemeyer,b Jeong-Gun Park,c Reena Allen,d Yoninah Cramer,c Stephen Murray,e Deborah Sutherland,f Francesca Aweeka,b Susan L. Koletar,g Florence Marzan,b Jing Bao,h Rada Savic,b David W. Haas,f the AIDS Clinical Trials Group A5306 Study Team Johns Hopkins University School of Medicine, Baltimore, Maryland, USAa; University of California, San Francisco, California, USAb; Statistical and Data Analysis Center, Harvard School of Public Health, Boston, Massachusetts, USAc; Social & Scientific Systems, Inc., Silver Spring, Maryland, USAd; Global Alliance for TB Drug Development, New York, New York, USAe; Vanderbilt University School of Medicine, Nashville, Tennessee, USAf; Ohio State University, Columbus, Ohio, USAg; Henry M. Jackson Foundation, Division of AIDS, National Institutes of Health, Bethesda, Maryland, USAh There is an urgent need for new antituberculosis (anti-TB) drugs, including agents that are safe and effective with concomitant antiretrovirals (ARV) and first-line TB drugs. PA-824 is a novel antituberculosis nitroimidazole in late-phase clinical develop- ment. Cytochrome P450 (CYP) 3A, which can be induced or inhibited by ARV and antituberculosis drugs, is a minor (ϳ20%) metabolic pathway for PA-824. In a phase I clinical trial, we characterized interactions between PA-824 and efavirenz (arm 1), lopinavir/ritonavir (arm 2), and rifampin (arm 3) in healthy, HIV-uninfected volunteers without TB disease. Participants in arms 1 and 2 were randomized to receive drugs via sequence 1 (PA-824 alone, washout, ARV, and ARV plus PA-824) or sequence 2 (ARV, ARV with PA-824, washout, and PA-824 alone). -

Medications to Be Avoided Or Used with Caution in Parkinson's Disease
Medications To Be Avoided Or Used With Caution in Parkinson’s Disease This medication list is not intended to be complete and additional brand names may be found for each medication. Every patient is different and you may need to take one of these medications despite caution against it. Please discuss your particular situation with your physician and do not stop any medication that you are currently taking without first seeking advice from your physician. Most medications should be tapered off and not stopped suddenly. Although you may not be taking these medications at home, one of these medications may be introduced while hospitalized. If a hospitalization is planned, please have your neurologist contact your treating physician in the hospital to advise which medications should be avoided. Medications to be avoided or used with caution in combination with Selegiline HCL (Eldepryl®, Deprenyl®, Zelapar®), Rasagiline (Azilect®) and Safinamide (Xadago®) Medication Type Medication Name Brand Name Narcotics/Analgesics Meperidine Demerol® Tramadol Ultram® Methadone Dolophine® Propoxyphene Darvon® Antidepressants St. John’s Wort Several Brands Muscle Relaxants Cyclobenzaprine Flexeril® Cough Suppressants Dextromethorphan Robitussin® products, other brands — found as an ingredient in various cough and cold medications Decongestants/Stimulants Pseudoephedrine Sudafed® products, other Phenylephrine brands — found as an ingredient Ephedrine in various cold and allergy medications Other medications Linezolid (antibiotic) Zyvox® that inhibit Monoamine oxidase Phenelzine Nardil® Tranylcypromine Parnate® Isocarboxazid Marplan® Note: Additional medications are cautioned against in people taking Monoamine oxidase inhibitors (MAOI), including other opioids (beyond what is mentioned in the chart above), most classes of antidepressants and other stimulants (beyond what is mentioned in the chart above). -

Using Medications, Cosmetics, Or Eating Certain Foods Can Increase Sensitivity to Ultraviolet Radiation
USING MEDICATIONS, COSMETICS, OR EATING CERTAIN FOODS CAN INCREASE SENSITIVITY TO ULTRAVIOLET RADIATION. INDIVIDUALS SHOULD CONSULT A PHYSICIAN BEFORE USING A SUNLAMP, IF THEY ARE TAKING MEDICATIONS. THE ITEMS LISTED ARE POTENTIAL PHOTOSENSITIZING AGENTS THAT MAY INCREASE SENSITIVITY TO ULTRAVIOLET LIGHT THAT MAY RESULT IN A PHOTOTOXIC OR PHOTOALLERGIC RESPONSES. PHOTOSENSITIZING MEDICATIONS Acetazolamide Amiloride+Hydrochlorothizide Amiodarone Amitriptyline Amoxapine Astemizole Atenolol+Chlorthalidone Auranofin Azatadine (Optimine) Azatidine+Pseudoephedrine Bendroflumethiazide Benzthiazide Bromodiphenhydramine Bromopheniramine Captopril Captopril+Hydrochlorothiazide Carbaamazepine Chlordiazepoxide+Amitriptyline Chlorothiazide Chlorpheniramine Chlorpheniramin+DPseudoephedrine Chlorpromazine Chlorpheniramine+Phenylopropanolamine Chlorpropamide Chlorprothixene Chlorthalidone Chlorthalidone+Reserpine Ciprofloxacin Clemastine Clofazime ClonidineChlorthalisone+Coal Tar Coal Tar Contraceptive (oral) Cyclobenzaprine Cyproheptadine Dacarcazine Danazol Demeclocycline Desipramine Dexchlorpheniramine Diclofenac Diflunisal Ditiazem Diphenhydramine Diphenylpyraline Doxepin Doxycycline Doxycycline Hyclate Enalapril Enalapril+Hydrochlorothiazide Erythromycin Ethylsuccinate+Sulfisoxazole Estrogens Estrogens Ethionamide Etretinate Floxuridine Flucytosine Fluorouracil Fluphenazine Flubiprofen Flutamide Gentamicin Glipizide Glyburide Gold Salts (compounds) Gold Sodium Thiomalate Griseofulvin Griseofulvin Ultramicrosize Griseofulvin+Hydrochlorothiazide Haloperidol -

Drug-Induced Diseases: Depression
Drug-Induced Diseases Section IV: Drug-Induced Psychiatric Diseases Chapter 18: Depression Sheila Botts and Melody Ryan Depression is one of the most common psychiatric illnesses, and it may interfere significantly with a patient’s daily functioning and quality of life. Untreated depression is associated with substantial morbidity and increases the risk of suicide and death. An estimated 15% of mood disorders end in suicide.1 Depression is a biologic illness with an unknown cause. Depression occurring secondary to medications is similar in presentation to endogenous depression and carries similar risks of morbidity and mortality. While the overall prevalence is unknown, drug-induced depression poses a significant challenge for practitioners, as it may undermine the effectiveness of much-needed treatment. The risk of treatment-emergent suicidality (e.g., suicidal ideation and behavior) has more recently been characterized independently, although it often occurs with mood changes and poses significant risks to the patient. Contents CAUSATIVE AGENTS ................................................................................................................. 1 EPIDEMIOLOGY .......................................................................................................................... 3 MECHANISMS .............................................................................................................................. 9 CLINICAL PRESENTATION AND DIFFERENTIAL DIAGNOSIS ........................................ 11 RISK FACTORS -

(Bedaquiline) with Efavirenz in Healthy Volunteers: AIDS Clinical Trials Group Study A5267
CLINICAL SCIENCE Safety, Tolerability, and Pharmacokinetic Interactions of the Antituberculous Agent TMC207 (Bedaquiline) With Efavirenz in Healthy Volunteers: AIDS Clinical Trials Group Study A5267 Kelly E. Dooley, MD, PhD,* Jeong-Gun Park, PhD,† Susan Swindells, MBBS,‡ Reena Allen, MA,§ David W. Haas, MD,¶ Yoninah Cramer, MS,† Francesca Aweeka, PharmD,# Ilene Wiggins, RN,* Amita Gupta, MD,* Patricia Lizak, BS,# Sonia Qasba, MD, MPH,†† Rolf van Heeswijk, PharmD, PhD,** Charles Flexner, MD,* and the ACTG 5267 Study Team Results: Thirty-three of 37 enrolled subjects completed the study. – Background: Drug drug interactions complicate management of Geometric mean of ratios for bedaquiline with efavirenz versus coinfection with HIV-1 and Mycobacterium tuberculosis. Bedaqui- bedaquiline alone were 0.82 [90% confidence interval (CI): 0.75 to line (formerly TMC207), an investigational agent for the treatment 0.89] for the 14-day area under the concentration–time curve of tuberculosis, is metabolized by cytochrome P450 (CYP) 3A (AUC0–336 h) and 1.00 (90% CI: 0.88 to 1.13) for the maximum which may be induced by the antiretroviral drug efavirenz. concentration (Cmax). For N-monodesmethyl metabolite, the geomet- Methods: This was a phase 1 pharmacokinetic drug interaction ric mean of ratios was 1.07 (90% CI: 0.97 to 1.19) for AUC0–336 h trial. Each healthy volunteer received two 400 mg doses of bedaqui- and 1.89 (90% CI: 1.66 to 2.15) for Cmax. There were no grade 3 or line, the first alone and the second with concomitant steady-state 4 clinical adverse events. One subject developed asymptomatic grade efavirenz. -

Anafranil (Clomipramine Hydrochloride) Tablets 1 Name of the Medicine 2 Qualitative and Quant
AUSTRALIAN PRODUCT INFORMATION – ANAFRANIL (CLOMIPRAMINE HYDROCHLORIDE) TABLETS 1 NAME OF THE MEDICINE Clomipramine hydrochloride 2 QUALITATIVE AND QUANTITATIVE COMPOSITION Each tablet contains 25 mg clomipramine hydrochloride. Lactose and sucrose Anafranil tablets contain lactose and sucrose. Patients with rare hereditary problems of galactose intolerance, fructose intolerance, severe lactase deficiency, sucrase-isomaltase insufficiency or glucose-galactose malabsorption should not take Anafranil tablets. For the full list of excipients, see Section 6.1 List of excipients. 3 PHARMACEUTICAL FORM Tablet 4 CLINICAL PARTICULARS 4.1 THERAPEUTIC INDICATIONS • Major depression. • Obsessive-compulsive disorders and phobias in adults. • Cataplexy associated with narcolepsy. 4.2 DOSE AND METHOD OF ADMINISTRATION General The dosage should be determined individually and adapted to the patient's condition. Doses should be kept as low as possible and increased cautiously. Note that the plasma concentrations of the drug and active metabolite do not stabilise for 7 to 14 days after commencing treatment and after a dosage change. During treatment, the efficacy and tolerability of Anafranil must be judged by keeping the patients under close surveillance. Depression, obsessive-compulsive disorders and phobias: Start treatment with one tablet of 25 mg 2 or 3 times daily. Raise the daily dosage stepwise, e.g. 25 mg every few days, (depending on how the medication is tolerated) to 4 to 6 tablets of 25 mg. Once a distinct improvement has set in, adjust the daily dosage to a maintenance level averaging 2 to 4 tablets of 25 mg. 1 Cataplexy accompanying narcolepsy: Anafranil should be given orally in a daily dose of 25 to 75 mg. -

Va / Dod Depression Practice Guideline Provider Care Card Antidepressant
VA / DOD DEPRESSION PRACTICE GUIDELINE PROVIDER CARE CARD CARD ANTIDEPRESSANT MEDICATION TABLE 7 Refer to pharmaceutical manufacturer’s literature for full prescribing information SEROTONIN SELECTIVE REUPTAKE INHIBITORS (SSRIs) GENERIC BRAND NAME ADULT STARTING DOSE MAX EXCEPTION SAFETY MARGIN TOLERABILITY EFFICACY SIMPLICITY Citalopram Celexa 20 mg 60 mg Reduce dose Nausea, insomnia, for the elderly & No serious systemic sedation, Fluoxetine Prozac 20 mg 80 mg those with renal toxicity even after headache, fatigue or hepatic substantial overdose. dizziness, sexual AM daily dosing. Paroxetine Paxil 20 mg 50 mg failure Drug interactions may dysfunction Response rate = Can be started at Sertraline Zoloft 50 mg 200 mg include tricyclic anorexia, weight 2 - 4 wks an effective dose antidepressants, loss, sweating, GI immediately. First Line Antidepressant Medication carbamazepine & distress, tremor, Drugs of this class differ substantially in safety, tolerability and simplicity when used in patients warfarin. restlessness, on other medications. Can work in TCA (tricyclic antidepressant) nonresponders. Useful in agitation, anxiety. several anxiety disorders. Taper gradually when discontinuing these medications. SEROTONIN and NOREPHINEPHRINE REUPTAKE INHIBITORS GENERIC BRAND NAME ADULT STARTING DOSE MAX EXCEPTION SAFETY MARGIN TOLERABILITY EFFICACY SIMPLICITY Reduce dose Take with food. Venlafaxine IR Effexor IR 75 mg 375 mg for the elderly & Comparable to BID or TID No serious systemic those with renal SSRIs at low dose. dosing with IR. toxicity. or hepatic Nausea, dry mouth, Response rate = Daily dosing Downtaper slowly to Venlafaxine XR Effexor XR 75 mg 375 mg failure insomnia, anxiety, 2 - 4 wks with XR. prevent clinically somnolence, head (4 - 7 days at Can be started at significant withdrawal ache, dizziness, ~300 mg/day) an effective dose Dual action drug that predominantly acts like a Serotonin Selective Reuptake inhibitor at low syndrome. -

Drug Induced Parkinsonism: an Overview
Review Article Open Access J Neurol Neurosurg Volume 3 Issue 4 - May 2017 Copyright © All rights are reserved by Sujith Ovallath DOI: 10.19080/OAJNN.2017.03.555620 Drug Induced Parkinsonism: An Overview Bahiya Sulthana and Sujith Ovallath* James Parkinson’s Research Centre, Kannur Medical College, India Submission: March 22, 2017; Published: May 05, 2017 *Corresponding author: Sujith Ovallath, James Parkinson’s Research Centre, Kannur Medical College, India, Email: Abstract Ever since 1950s from reserpine rat model to [18F] FP-CIT PET scan, our knowledge of drug induced Parkinsonism has been constantly evolving. Trend of suspected agents changed from typical antipsychotic to more commonly used drugs like Metoclopramide and more recently to newer prokinetics like levosulpiride. Even though DIP is mostly reversible, unmasking of subclinical PD and irreversible damage by the drug leads to persistent DIP. Demonstrated risk is higher with elderly and women. Symptoms can appear from 3 months (69%) to 12 months (20%) of the drug intake. Most of causative agents act by blocking the dopaminergic transmission in niagrostriatal pathway. The clinical syndrome is likely to present with subacute onset with symmetrical involvement. Presence of non-motor symptoms especially hyposmia and asymmetrical involvement is suggestive of degenerative Parkinsonism. DaT scan/SPECT are the investigation available to differentiate pure DIP from other forms. Management of this condition comprises of recognition of symptoms early identifying the offending agents and prompt discontinuation of the causal agent . This result in marked improvement in about 90% patients. Medical therapy is indicated only if the symptoms persist or are problematic. Symptomatic treatment includes anti-cholinergic drugs like benztropine and trihexifenidyl, amantidine and ECT for refractory cases. -

Pharmacological Drugs Inducing Ototoxicity, Vestibular Symptoms and Tinnitus: a Reasoned and Updated Guide
European Review for Medical and Pharmacological Sciences 2011; 15: 601-636 Pharmacological drugs inducing ototoxicity, vestibular symptoms and tinnitus: a reasoned and updated guide G. CIANFRONE1, D. PENTANGELO1, F. CIANFRONE2, F. MAZZEI1, R. TURCHETTA1, M.P. ORLANDO1, G. ALTISSIMI1 1Department of Otolaryngology, Audiology and Phoniatrics, “Umberto I” University Hospital, Sapienza University, Rome (Italy); 2Institute of Otorhinolaryngology, School of Medicine, Catholic University of the Sacred Heart, Rome (Italy) Abstract. – The present work on drug-in- Introduction duced ototoxicity, tinnitus and vertigo repre- sents the update and revision of a previous The panorama of the pharmacological origin guide to adverse drug reactions for italian physi- iatrogenic noxae able to induce either harmful cians (2005). The panorama of drug-induced side effects causing ototoxicity or symptoms ototoxic effects or just a symptomatology like tin- such as tinnitus or dizziness and vertigo has en- nitus or balance disturbances, without any harm- larged in recent years, thanks to a better knowl- ful consequence, has widened in the last few edge and a more specific attention of pharma- years. The reason for this is the progress of scien- ceutical firms and drug-control institutions. In tific knowledge, the increased awareness of the daily clinical practice, there is a need for the pharmaceutical companies and of the institutions, family physician and the ENT specialist or audi- ologist (also in consideration of the possible which supervise pharmaceutical production. medico-legal implications) to focus the attention Only through continuous updating and experi- on the possible risk of otological side effects. ence sharing it’s possible to offer patients the This would allow a clinical risk-benefit evalua- certainty of receiving the treatment that is appro- tion, weighing the possible clinical advantage in priate, safe and effective and based upon the their field of competence against possible oto- most credited clinical studies.