Xerox University Microfilms
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ostrich Production Systems Part I: a Review
11111111111,- 1SSN 0254-6019 Ostrich production systems Food and Agriculture Organization of 111160mmi the United Natiorp str. ro ucti s ct1rns Part A review by Dr M.M. ,,hanawany International Consultant Part II Case studies by Dr John Dingle FAO Visiting Scientist Food and , Agriculture Organization of the ' United , Nations Ot,i1 The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. M-21 ISBN 92-5-104300-0 Reproduction of this publication for educational or other non-commercial purposes is authorized without any prior written permission from the copyright holders provided the source is fully acknowledged. Reproduction of this publication for resale or other commercial purposes is prohibited without written permission of the copyright holders. Applications for such permission, with a statement of the purpose and extent of the reproduction, should be addressed to the Director, Information Division, Food and Agriculture Organization of the United Nations, Viale dells Terme di Caracalla, 00100 Rome, Italy. C) FAO 1999 Contents PART I - PRODUCTION SYSTEMS INTRODUCTION Chapter 1 ORIGIN AND EVOLUTION OF THE OSTRICH 5 Classification of the ostrich in the animal kingdom 5 Geographical distribution of ratites 8 Ostrich subspecies 10 The North -

Estimating Mass Properties of Dinosaurs Using Laser Imaging and 3D Computer Modelling
Estimating Mass Properties of Dinosaurs Using Laser Imaging and 3D Computer Modelling Karl T. Bates1*, Phillip L. Manning2,3, David Hodgetts3, William I. Sellers1 1 Adaptive Organismal Biology Research Group, Faculty of Life Sciences, University of Manchester, Jackson’s Mill, Manchester, United Kingdom, 2 The Manchester Museum, University of Manchester, Manchester, United Kingdom, 3 School of Earth, Atmospheric and Environmental Science, University of Manchester, Manchester, United Kingdom Abstract Body mass reconstructions of extinct vertebrates are most robust when complete to near-complete skeletons allow the reconstruction of either physical or digital models. Digital models are most efficient in terms of time and cost, and provide the facility to infinitely modify model properties non-destructively, such that sensitivity analyses can be conducted to quantify the effect of the many unknown parameters involved in reconstructions of extinct animals. In this study we use laser scanning (LiDAR) and computer modelling methods to create a range of 3D mass models of five specimens of non- avian dinosaur; two near-complete specimens of Tyrannosaurus rex, the most complete specimens of Acrocanthosaurus atokensis and Strutiomimum sedens, and a near-complete skeleton of a sub-adult Edmontosaurus annectens. LiDAR scanning allows a full mounted skeleton to be imaged resulting in a detailed 3D model in which each bone retains its spatial position and articulation. This provides a high resolution skeletal framework around which the body cavity and internal organs such as lungs and air sacs can be reconstructed. This has allowed calculation of body segment masses, centres of mass and moments or inertia for each animal. However, any soft tissue reconstruction of an extinct taxon inevitably represents a best estimate model with an unknown level of accuracy. -

Postcranial Skeletal Pneumaticity in Sauropods and Its
Postcranial Pneumaticity in Dinosaurs and the Origin of the Avian Lung by Mathew John Wedel B.S. (University of Oklahoma) 1997 A dissertation submitted in partial satisfaction of the requirements for the degree of Doctor of Philosophy in Integrative Biology in the Graduate Division of the University of California, Berkeley Committee in charge: Professor Kevin Padian, Co-chair Professor William Clemens, Co-chair Professor Marvalee Wake Professor David Wake Professor John Gerhart Spring 2007 1 The dissertation of Mathew John Wedel is approved: Co-chair Date Co-chair Date Date Date Date University of California, Berkeley Spring 2007 2 Postcranial Pneumaticity in Dinosaurs and the Origin of the Avian Lung © 2007 by Mathew John Wedel 3 Abstract Postcranial Pneumaticity in Dinosaurs and the Origin of the Avian Lung by Mathew John Wedel Doctor of Philosophy in Integrative Biology University of California, Berkeley Professor Kevin Padian, Co-chair Professor William Clemens, Co-chair Among extant vertebrates, postcranial skeletal pneumaticity is present only in birds. In birds, diverticula of the lungs and air sacs pneumatize specific regions of the postcranial skeleton. The relationships among pulmonary components and the regions of the skeleton that they pneumatize form the basis for inferences about the pulmonary anatomy of non-avian dinosaurs. Fossae, foramina and chambers in the postcranial skeletons of pterosaurs and saurischian dinosaurs are diagnostic for pneumaticity. In basal saurischians only the cervical skeleton is pneumatized. Pneumatization by cervical air sacs is the most consilient explanation for this pattern. In more derived sauropods and theropods pneumatization of the posterior dorsal, sacral, and caudal vertebrae indicates that abdominal air sacs were also present. -

'Big Al'? Quantifying the Effect of Soft
Palaeontologia Electronica http://palaeo-electronica.org HOW BIG WAS ‘BIG AL’? QUANTIFYING THE EFFECT OF SOFT TISSUE AND OSTEOLOGICAL UNKNOWNS ON MASS PREDICTIONS FOR ALLOSAURUS (DINOSAURIA:THEROPODA) Karl T. Bates, Peter L. Falkingham, Brent H. Breithaupt, David Hodgetts, William I. Sellers, and Phillip L. Manning ABSTRACT MOR693, nicknamed ‘Big Al,’ is the most complete skeleton of the non-avian theropod Allosaurus and therefore provides the best opportunity to investigate the mass properties of this important Jurassic theropod through accurate physical or digital volumetric models. In this study, laser scanning and computer modelling software have been used to construct volumetric models of MOR693. A long-range laser scanner has been used to digitize the mounted cast of MOR693, allowing the reconstruction of body volumes and respiratory structures around and within the three-dimensional (3D) skel- etal model. The digital medium offered the facility to modify model properties non- destructively in a detailed sensitivity analysis to quantify the effects of the many unknown parameters involved in such reconstructions. In addition to varying the vol- umes of body segments and respiratory structures, we also extend the sensitivity anal- ysis to include uncertainties regarding osteological articulations in non-avian dinosaurs, including effects of inter-vertebral spacing and the orientation or ‘flare’ of the rib cage in MOR693. Results suggest body mass and inertial values are extremely uncertain and show a wide range in plausible values, whilst the CM (centre of mass) position is well constrained immediately in front and below the hip joint in MOR693, consistent with similar reconstructions of non-avian theropods. Karl T. -
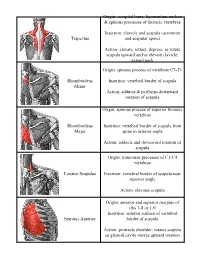
Trapezius Origin: Occipital Bone, Ligamentum Nuchae & Spinous Processes of Thoracic Vertebrae Insertion: Clavicle and Scapul
Origin: occipital bone, ligamentum nuchae & spinous processes of thoracic vertebrae Insertion: clavicle and scapula (acromion Trapezius and scapular spine) Action: elevate, retract, depress, or rotate scapula upward and/or elevate clavicle; extend neck Origin: spinous process of vertebrae C7-T1 Rhomboideus Insertion: vertebral border of scapula Minor Action: adducts & performs downward rotation of scapula Origin: spinous process of superior thoracic vertebrae Rhomboideus Insertion: vertebral border of scapula from Major spine to inferior angle Action: adducts and downward rotation of scapula Origin: transverse precesses of C1-C4 vertebrae Levator Scapulae Insertion: vertebral border of scapula near superior angle Action: elevates scapula Origin: anterior and superior margins of ribs 1-8 or 1-9 Insertion: anterior surface of vertebral Serratus Anterior border of scapula Action: protracts shoulder: rotates scapula so glenoid cavity moves upward rotation Origin: anterior surfaces and superior margins of ribs 3-5 Insertion: coracoid process of scapula Pectoralis Minor Action: depresses & protracts shoulder, rotates scapula (glenoid cavity rotates downward), elevates ribs Origin: supraspinous fossa of scapula Supraspinatus Insertion: greater tuberacle of humerus Action: abduction at the shoulder Origin: infraspinous fossa of scapula Infraspinatus Insertion: greater tubercle of humerus Action: lateral rotation at shoulder Origin: clavicle and scapula (acromion and adjacent scapular spine) Insertion: deltoid tuberosity of humerus Deltoid Action: -

Coracoid Process Anatomy: a Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B
Quantitative Assessment of the Coracoacromial and the Coracoclavicular Ligaments With 3-Dimensional Mapping of the Coracoid Process Anatomy: A Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B. Ferrari, M.D., George Sanchez, B.S., Alex W. Brady, M.Sc., Jonas Pogorzelski, M.D., M.H.B.A., George F. Lebus, M.D., Peter J. Millett, M.D., M.Sc., Robert F. LaPrade, M.D., Ph.D., and CAPT Matthew T. Provencher, M.D., M.C., U.S.N.R. Purpose: To perform a quantitative anatomic evaluation of the (1) coracoid process, specifically the attachment sites of the conjoint tendon, the pectoralis minor, the coracoacromial ligament (CAL), and the coracoclavicular (CC) ligaments in relation to pertinent osseous and soft tissue landmarks; (2) CC ligaments’ attachments on the clavicle; and (3) CAL attachment on the acromion in relation to surgically relevant anatomic landmarks to assist in planning of the Latarjet procedure, acromioclavicular (AC) joint reconstructions, and CAL resection distances avoiding iatrogenic injury to sur- rounding structures. Methods: Ten nonpaired fresh-frozen human cadaveric shoulders (mean age 52 years, range 33- 64 years) were included in this study. A 3-dimensional coordinate measuring device was used to quantify the location of pertinent bony landmarks and soft tissue attachment areas. The ligament and tendon attachment perimeters and center points on the coracoid, clavicle, and acromion were identified and subsequently dissected off the bone. Coordinates of points along the perimeters of attachment sites were used to calculate areas, whereas coordinates of center points were used to determine distances between surgically relevant attachment sites and pertinent bony landmarks. -

Scientific American
JANUARY 2021 SCIENTIFICAMERICAN.COM How COVID-19 Wrecks the Immune System Ancient Galaxy Clusters Understanding Mountain Ice DINO STAR Scientists reveal the real Dilophosaurus, a Jurassic Park icon © 2020 Scientific American JANUARY 2021 VOLUME 324, NUMBER 1 0068 A S TRONOMY PALEONTOLOGY 26 Too Big for the Universe 46 The Real Dilophosaurus Ancient galaxy clusters seem to The most comprehensive study have grown so quickly that they of the iconic Jurassic Park dinosaur would have broken the laws reveals a very different animal of the cosmos. By Arianna S. Long from the one the movie portrayed. By Matthew A. Brown and IMMUNOLOGY Adam D. Marsh 34 The Immune Havoc of COVID-19 NATURAL RESOURCES The virus flourishes by under 54 Peak Water mining the body’s chemical Data retrieved from Earth’s highest defense system. mountains show that the water By Akiko Iwasaki and supply to two billion people Patrick Wong is changing. By Walter Immerzeel PUBLIC HEALTH MEDICINE 42 The Very Real Death Toll 62 Malignant Cheaters of COVID-19 Cells coexist by cooperating. President Trump and other When some break the rules, conspiracy fantasists touted cancers result. By Athena Aktipis ON THE COVER Jurassic Park made Dilophosaurus famous before the fake claim that COVID death SPACE SCIENCE scientists had a thorough understanding of counts are exaggerated. But 68 Dynamic Planet this dinosaur. A new analysis of Dilophosaurus three kinds of evidence point For 15 years the Mars Reconnais remains has provided the most detailed picture to more than 250,000 deaths, sance Orbiter has transformed yet of a dinosaur of its vintage and revealed the creature as it truly was: a large-bodied, nimble a toll that grows every day. -

Reconstructing Pectoral Appendicular Muscle Anatomy in Fossil Fish and Tetrapods Over the Fins-To-Limbs Transition
Biol. Rev. (2017), pp. 000–000. 1 doi: 10.1111/brv.12386 Reconstructing pectoral appendicular muscle anatomy in fossil fish and tetrapods over the fins-to-limbs transition Julia L. Molnar1,∗ , Rui Diogo2, John R. Hutchinson3 and Stephanie E. Pierce4 1Department of Anatomy, New York Institute of Technology College of Osteopathic Medicine, Northern Boulevard, Old Westbury, NY, U.S.A. 2Department of Anatomy, Howard University College of Medicine, 520 W St. NW, Numa Adams Building, Washington, DC 20059, U.S.A. 3Structure and Motion Lab, Royal Veterinary College, Hawkshead Lane, Hatfield, Hertfordshire AL9 7TA, UK 4Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, U.S.A. ABSTRACT The question of how tetrapod limbs evolved from fins is one of the great puzzles of evolutionary biology. While palaeontologists, developmental biologists, and geneticists have made great strides in explaining the origin and early evolution of limb skeletal structures, that of the muscles remains largely unknown. The main reason is the lack of consensus about appendicular muscle homology between the closest living relatives of early tetrapods: lobe-finned fish and crown tetrapods. In the light of a recent study of these homologies, we re-examined osteological correlates of muscle attachment in the pectoral girdle, humerus, radius, and ulna of early tetrapods and their close relatives. Twenty-nine extinct and six extant sarcopterygians were included in a meta-analysis using information from the literature and from original specimens, when possible. We analysed these osteological correlates using parsimony-based character optimization in order to reconstruct muscle anatomy in ancestral lobe-finned fish, tetrapodomorph fish, stem tetrapods, and crown tetrapods. -

Breathing and Locomotion in Birds
Breathing and locomotion in birds. A thesis submitted to the University of Manchester for the degree of Doctor of Philosophy in the Faculty of Life Sciences. 2010 Peter George Tickle Contents Abstract 4 Declaration 5 Copyright Statement 6 Author Information 7 Acknowledgements 9 Organisation of this PhD thesis 10 Chapter 1 General Introduction 13 1. Introduction 14 1.1 The Avian Respiratory System 14 1.1.1 Structure of the lung and air sacs 16 1.1.2 Airflow in the avian respiratory system 21 1.1.3 The avian aspiration pump 25 1.2 The uncinate processes in birds 29 1.2.1 Uncinate process morphology and biomechanics 32 1.3 Constraints on breathing in birds 33 1.3.1 Development 33 1.3.2 Locomotion 35 1.3.2.1 The appendicular skeleton 35 1.3.2.2 Overcoming the trade-off between breathing 36 and locomotion 1.3.2.3 Energetics of locomotion in birds 38 1.4 Evolution of the ventilatory pump in birds 41 1.5 Overview and Thesis Aims 42 2 Chapter 2 Functional significance of the uncinate processes in birds. 44 Chapter 3 Ontogenetic development of the uncinate processes in the 45 domestic turkey (Meleagris gallopavo). Chapter 4 Uncinate process length in birds scales with resting metabolic rate. 46 Chapter 5 Load carrying during locomotion in the barnacle goose (Branta 47 leucopsis): The effect of load placement and size. Chapter 6 A continuum in ventilatory mechanics from early theropods to 48 extant birds. Chapter 7 General Discussion 49 References 64 3 Abstract of a thesis by Peter George Tickle submitted to the University of Manchester for the degree of PhD in the Faculty of Life Sciences and entitled ‘Breathing and Locomotion in Birds’. -

Wedel, M. J., & Taylor, M. P. (2013). Caudal Pneumaticity And
Wedel, M. J. , & Taylor, M. P. (2013). Caudal pneumaticity and pneumatic hiatuses in the sauropod dinosaurs Giraffatitan and Apatosaurus. PLoS ONE, 8(10), [e78213]. https://doi.org/10.1371/journal.pone.0078213 Publisher's PDF, also known as Version of record Link to published version (if available): 10.1371/journal.pone.0078213 Link to publication record in Explore Bristol Research PDF-document CC By University of Bristol - Explore Bristol Research General rights This document is made available in accordance with publisher policies. Please cite only the published version using the reference above. Full terms of use are available: http://www.bristol.ac.uk/red/research-policy/pure/user-guides/ebr-terms/ Caudal Pneumaticity and Pneumatic Hiatuses in the Sauropod Dinosaurs Giraffatitan and Apatosaurus Mathew J. Wedel1*, Michael P. Taylor2* 1 College of Osteopathic Medicine of the Pacific and College of Podiatric Medicine, Western University of Health Sciences, Pomona, California, United States of America, 2 Department of Earth Sciences, University of Bristol, United Kingdom Abstract Skeletal pneumaticity is found in the presacral vertebrae of most sauropod dinosaurs, but pneumaticity is much less common in the vertebrae of the tail. We describe previously unrecognized pneumatic fossae in the mid-caudal vertebrae of specimens of Giraffatitan and Apatosaurus. In both taxa, the most distal pneumatic vertebrae are separated from other pneumatic vertebrae by sequences of three to seven apneumatic vertebrae. Caudal pneumaticity is not prominent in most individuals of either of these taxa, and its unpredictable development means that it may be more widespread than previously recognised within Sauropoda and elsewhere in Saurischia. -

Some Notes on the Diverse Brachiosaurid Sauropods of the Late Jurassic of North America, Europe and Africa
Some Notes on the Diverse Brachiosaurid Sauropods of the Late Jurassic of North America, Europe and Africa GREGORY S. PAUL 3109 N. Calvert St. Baltimore, Maryland 21218 USA [email protected] June 2012 Abstract: The unusual placement of the parapophyses on elongotated peduncles further distinguishes Brachiosaurus from other brachiosaurid genera including Giraffatitan. Late Jurassic brachiosaurid taxonomy is probably more complex than has been realized, in part because of evolution over the considerable periods of times recorded in the Morrison and Tendaguru formations. The presence of reduced tails on late Jurassic brachiosaurids is confirmed. INTRODUCTION It was long assumed that the brachiosaurid material found in the Late Jurassic Morrison, Tendaguru, and Lourinha Formations from three continents belonged to the genus Brachiosaurus (as per Janensch 1950, 1961, Lapparent and Zbyszewski 1957, Jensen 1987) until Paul (1988) noted significant differences between the type species B. altithorax (Fig. 1A) from North America and the African B. brancai (Fig. 1B) indicated at least a subgeneric separation, and Taylor (2009) formally separated the sauropods, leaving much of the Tendaguru brachiosaurids material titled Giraffatitan brancai; the generic separation has been tentatively questioned (Chure et al. 2010). In addition European B. alatalaiensis has been retitled Lusotitan alatalaiensis (Antunes and Mateus 2003), and a dwarf brachiosaur from the European Mittlere Kimmeridge-Stufe deposits has been designated Europasaurus holgeri (Sander et al. 2006). Institutional Abbreviations: AMNH, American Museum of Natural History, New York; BYU, Brigham Young University, Provo; FMNH, Field Museum of Natural History, Chicago; HMN, Humboldt Museum fur Naturkunde, Berlin; NHMUK, Natural History Museum, London; USNM, United States Natural History Museum, Washington DC. -

Rehabilitation Guidelines for Latarjet/Coracoid Process Transfer Josef K
Rehabilitation Guidelines for Latarjet/Coracoid Process Transfer Josef K. Eichinger, MD Shoulder instability may be caused from congenital deformity, recurrent overuse activity, and/or traumatic dislocation. Surgical stabilization of the glenohumeral joint is indicated after conservative treatment fails and recurrent instability/subluxation continues. A number of different surgical procedures may be indicated in this situation, often divided into soft tissue or bony procedures. Shoulder Instability – Soft Tissue: Surgical reconstruction targeting the glenohumeral joint’s soft tissues for shoulder instability, typically involves labral repairs, the most common being the Bankart repair. A Bankart lesion typically occurs from an anterior-inferior dislocation of the humerus, tearing the labrum from it’s attachment to the glenoid, thereby detaching the inferior gleno-humeral ligament (IGHL). Surgical management of this revolves around labral repair to reattach the IGHL under appropriate tension. This may be accomplished either arthroscopically or through an open approach.1 Most traumatic glenohumeral dislocations may not only cause a Bankart lesion, but may create impression fractures in the postero-superior humeral head termed Hill-Sachs lesions.2 An adverse effect from this procedure includes suturing the capsule too tightly, causing a shortening of the capsule, and thus decreasing the external rotation allowed at the glenohumeral joint. Other complications are extremely rare, but may include axillary nerve damage, subscapularis rupture (seen only in open repairs performed with subscapularis detachment and repair), and recurrent instability. If there is bony deficiency in the glenoid, which represents 20% or more of the antero-inferior glenoid, it is biomechanically impossible to restore the same stability and is therefore more prone to recurrent instability and failure.