DEINOCHEIRIDAE, a NEW FAMILY of THEROPOD DINOSAURS (Plates I-V)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fused and Vaulted Nasals of Tyrannosaurid Dinosaurs: Implications for Cranial Strength and Feeding Mechanics
Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for cranial strength and feeding mechanics ERIC SNIVELY, DONALD M. HENDERSON, and DOUG S. PHILLIPS Snively, E., Henderson, D.M., and Phillips, D.S. 2006. Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for cranial strength and feeding mechanics. Acta Palaeontologica Polonica 51 (3): 435–454. Tyrannosaurid theropods display several unusual adaptations of the skulls and teeth. Their nasals are fused and vaulted, suggesting that these elements braced the cranium against high feeding forces. Exceptionally high strengths of maxillary teeth in Tyrannosaurus rex indicate that it could exert relatively greater feeding forces than other tyrannosaurids. Areas and second moments of area of the nasals, calculated from CT cross−sections, show higher nasal strengths for large tyrannosaurids than for Allosaurus fragilis. Cross−sectional geometry of theropod crania reveals high second moments of area in tyrannosaurids, with resulting high strengths in bending and torsion, when compared with the crania of similarly sized theropods. In tyrannosaurids trends of strength increase are positively allomeric and have similar allometric expo− nents, indicating correlated progression towards unusually high strengths of the feeding apparatus. Fused, arched nasals and broad crania of tyrannosaurids are consistent with deep bites that impacted bone and powerful lateral movements of the head for dismembering prey. Key words: Theropoda, Carnosauria, Tyrannosauridae, biomechanics, feeding mechanics, computer modeling, com− puted tomography. Eric Snively [[email protected]], Department of Biological Sciences, University of Calgary, 2500 University Drive NW, Calgary, Alberta T2N 1N4, Canada; Donald M. Henderson [[email protected]], Royal Tyrrell Museum of Palaeontology, Box 7500, Drumheller, Alberta T0J 0Y0, Canada; Doug S. -

Evidence of a New Carcharodontosaurid from the Upper Cretaceous of Morocco
http://app.pan.pl/SOM/app57-Cau_etal_SOM.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Evidence of a new carcharodontosaurid from the Upper Cretaceous of Morocco Andrea Cau, Fabio Marco Dalla Vecchia, and Matteo Fabbri Published in Acta Palaeontologica Polonica 2012 57 (3): 661-665. http://dx.doi.org/10.4202/app.2011.0043 SOM1. PHYLOGENETIC ANALYSIS Material and Methods The data set of the phylogenetic analysis includes 37 Operational Taxonomic Units (OTUs) (35 ingroup neotheropod taxa, including one based on MPM 2594; and two basal theropod outgroups, Herrerasaurus and Tawa), and 808 character statements (see S2 and S3, below). The phylogenetic analyses were conducted through PAUP* vers. 4.010b (Swofford 2002). In the analysis, Herrerasaurus was used as root of the tree. The tree search strategy follows Benson (2009). The analysis initially performed a preliminary search using PAUPRat (Sikes and Lewis 2001). The unique topologies among the most parsimonious trees (MPTs) resulted after the preliminary search were used as a starting point for 1000 tree-bisection-reconnection branch swapping heuristic searches using PAUP*. Included taxa (and source for codings) Outgroup Herrerasaurus (Sereno 1993; Sereno and Novas 1993; Novas 1994; Sereno 2007) Tawa (Nesbitt et al. 2009) Ingroup Abelisaurus (Bonaparte and Novas 1985; Carrano and Sampson 2008) Acrocanthosaurus (Stoval and Landston 1950; Harris 1998; Currie and Carpenter 2000; Eddy and Clarke 2011) Allosaurus (Gilmore 1920; Madsen 1976) Baryonyx (Charig and Milner 1997; Mateus et al. 2010) Carcharodontosaurus (Stromer 1931; Sereno et al. 1996; Brusatte and Sereno 2007) Carnotaurus (Bonaparte et al. 1990) Ceratosaurus (Gilmore 1920; Madsen and Welles 2000). Compsognathus (Peyer 2006) Cryolophosaurus (Smith et al. -

The Origin and Early Evolution of Dinosaurs
Biol. Rev. (2010), 85, pp. 55–110. 55 doi:10.1111/j.1469-185X.2009.00094.x The origin and early evolution of dinosaurs Max C. Langer1∗,MartinD.Ezcurra2, Jonathas S. Bittencourt1 and Fernando E. Novas2,3 1Departamento de Biologia, FFCLRP, Universidade de S˜ao Paulo; Av. Bandeirantes 3900, Ribeir˜ao Preto-SP, Brazil 2Laboratorio de Anatomia Comparada y Evoluci´on de los Vertebrados, Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Avda. Angel Gallardo 470, Cdad. de Buenos Aires, Argentina 3CONICET (Consejo Nacional de Investigaciones Cient´ıficas y T´ecnicas); Avda. Rivadavia 1917 - Cdad. de Buenos Aires, Argentina (Received 28 November 2008; revised 09 July 2009; accepted 14 July 2009) ABSTRACT The oldest unequivocal records of Dinosauria were unearthed from Late Triassic rocks (approximately 230 Ma) accumulated over extensional rift basins in southwestern Pangea. The better known of these are Herrerasaurus ischigualastensis, Pisanosaurus mertii, Eoraptor lunensis,andPanphagia protos from the Ischigualasto Formation, Argentina, and Staurikosaurus pricei and Saturnalia tupiniquim from the Santa Maria Formation, Brazil. No uncontroversial dinosaur body fossils are known from older strata, but the Middle Triassic origin of the lineage may be inferred from both the footprint record and its sister-group relation to Ladinian basal dinosauromorphs. These include the typical Marasuchus lilloensis, more basal forms such as Lagerpeton and Dromomeron, as well as silesaurids: a possibly monophyletic group composed of Mid-Late Triassic forms that may represent immediate sister taxa to dinosaurs. The first phylogenetic definition to fit the current understanding of Dinosauria as a node-based taxon solely composed of mutually exclusive Saurischia and Ornithischia was given as ‘‘all descendants of the most recent common ancestor of birds and Triceratops’’. -

Estimating Mass Properties of Dinosaurs Using Laser Imaging and 3D Computer Modelling
Estimating Mass Properties of Dinosaurs Using Laser Imaging and 3D Computer Modelling Karl T. Bates1*, Phillip L. Manning2,3, David Hodgetts3, William I. Sellers1 1 Adaptive Organismal Biology Research Group, Faculty of Life Sciences, University of Manchester, Jackson’s Mill, Manchester, United Kingdom, 2 The Manchester Museum, University of Manchester, Manchester, United Kingdom, 3 School of Earth, Atmospheric and Environmental Science, University of Manchester, Manchester, United Kingdom Abstract Body mass reconstructions of extinct vertebrates are most robust when complete to near-complete skeletons allow the reconstruction of either physical or digital models. Digital models are most efficient in terms of time and cost, and provide the facility to infinitely modify model properties non-destructively, such that sensitivity analyses can be conducted to quantify the effect of the many unknown parameters involved in reconstructions of extinct animals. In this study we use laser scanning (LiDAR) and computer modelling methods to create a range of 3D mass models of five specimens of non- avian dinosaur; two near-complete specimens of Tyrannosaurus rex, the most complete specimens of Acrocanthosaurus atokensis and Strutiomimum sedens, and a near-complete skeleton of a sub-adult Edmontosaurus annectens. LiDAR scanning allows a full mounted skeleton to be imaged resulting in a detailed 3D model in which each bone retains its spatial position and articulation. This provides a high resolution skeletal framework around which the body cavity and internal organs such as lungs and air sacs can be reconstructed. This has allowed calculation of body segment masses, centres of mass and moments or inertia for each animal. However, any soft tissue reconstruction of an extinct taxon inevitably represents a best estimate model with an unknown level of accuracy. -

Xjiiie'icanj/Useum
XJiiie'ican1ox4tatreJ/useum PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK 24, N.Y. NUMBER 2I8I JUNE 4, I964 Relationships of the Saurischian Dinosaurs BY EDWIN H. COLBERT1 INTRODUCTION The word "Dinosauria" was coined by Sir Richard Owen in 1842 as a designation for various genera and species of extinct reptiles, the fossil bones of which were then being discovered and described in Europe. For many years this term persisted as the name for one order of reptiles and thus became well intrenched within the literature of paleontology. In- deed, since this name was associated with fossil remains that are frequently of large dimensions and spectacular shape and therefore of considerable interest to the general public, it in time became Anglicized, to take its proper place as a common noun in the English language. Almost every- body in the world is today more or less familiar with dinosaurs. As long ago as 1888, H. G. Seeley recognized the fact that the dino- saurs are not contained within a single reptilian order, but rather are quite clearly members of two distinct orders, each of which can be de- fined on the basis of many osteological characters. The structure of the pelvis is particularly useful in the separation of the two dinosaurian orders, and consequently Seeley named these two major taxonomic categories the Saurischia and the Ornithischia. This astute observation by Seeley was not readily accepted, so that for many years following the publication of his original paper proposing the basic dichotomy of the dinosaurs the 1 Chairman and Curator, Department ofVertebrate Paleontology, the American Museum of Natural History. -

Dinosaurs British Isles
DINOSAURS of the BRITISH ISLES Dean R. Lomax & Nobumichi Tamura Foreword by Dr Paul M. Barrett (Natural History Museum, London) Skeletal reconstructions by Scott Hartman, Jaime A. Headden & Gregory S. Paul Life and scene reconstructions by Nobumichi Tamura & James McKay CONTENTS Foreword by Dr Paul M. Barrett.............................................................................10 Foreword by the authors........................................................................................11 Acknowledgements................................................................................................12 Museum and institutional abbreviations...............................................................13 Introduction: An age-old interest..........................................................................16 What is a dinosaur?................................................................................................18 The question of birds and the ‘extinction’ of the dinosaurs..................................25 The age of dinosaurs..............................................................................................30 Taxonomy: The naming of species.......................................................................34 Dinosaur classification...........................................................................................37 Saurischian dinosaurs............................................................................................39 Theropoda............................................................................................................39 -

Postcranial Skeletal Pneumaticity in Sauropods and Its
Postcranial Pneumaticity in Dinosaurs and the Origin of the Avian Lung by Mathew John Wedel B.S. (University of Oklahoma) 1997 A dissertation submitted in partial satisfaction of the requirements for the degree of Doctor of Philosophy in Integrative Biology in the Graduate Division of the University of California, Berkeley Committee in charge: Professor Kevin Padian, Co-chair Professor William Clemens, Co-chair Professor Marvalee Wake Professor David Wake Professor John Gerhart Spring 2007 1 The dissertation of Mathew John Wedel is approved: Co-chair Date Co-chair Date Date Date Date University of California, Berkeley Spring 2007 2 Postcranial Pneumaticity in Dinosaurs and the Origin of the Avian Lung © 2007 by Mathew John Wedel 3 Abstract Postcranial Pneumaticity in Dinosaurs and the Origin of the Avian Lung by Mathew John Wedel Doctor of Philosophy in Integrative Biology University of California, Berkeley Professor Kevin Padian, Co-chair Professor William Clemens, Co-chair Among extant vertebrates, postcranial skeletal pneumaticity is present only in birds. In birds, diverticula of the lungs and air sacs pneumatize specific regions of the postcranial skeleton. The relationships among pulmonary components and the regions of the skeleton that they pneumatize form the basis for inferences about the pulmonary anatomy of non-avian dinosaurs. Fossae, foramina and chambers in the postcranial skeletons of pterosaurs and saurischian dinosaurs are diagnostic for pneumaticity. In basal saurischians only the cervical skeleton is pneumatized. Pneumatization by cervical air sacs is the most consilient explanation for this pattern. In more derived sauropods and theropods pneumatization of the posterior dorsal, sacral, and caudal vertebrae indicates that abdominal air sacs were also present. -

'Big Al'? Quantifying the Effect of Soft
Palaeontologia Electronica http://palaeo-electronica.org HOW BIG WAS ‘BIG AL’? QUANTIFYING THE EFFECT OF SOFT TISSUE AND OSTEOLOGICAL UNKNOWNS ON MASS PREDICTIONS FOR ALLOSAURUS (DINOSAURIA:THEROPODA) Karl T. Bates, Peter L. Falkingham, Brent H. Breithaupt, David Hodgetts, William I. Sellers, and Phillip L. Manning ABSTRACT MOR693, nicknamed ‘Big Al,’ is the most complete skeleton of the non-avian theropod Allosaurus and therefore provides the best opportunity to investigate the mass properties of this important Jurassic theropod through accurate physical or digital volumetric models. In this study, laser scanning and computer modelling software have been used to construct volumetric models of MOR693. A long-range laser scanner has been used to digitize the mounted cast of MOR693, allowing the reconstruction of body volumes and respiratory structures around and within the three-dimensional (3D) skel- etal model. The digital medium offered the facility to modify model properties non- destructively in a detailed sensitivity analysis to quantify the effects of the many unknown parameters involved in such reconstructions. In addition to varying the vol- umes of body segments and respiratory structures, we also extend the sensitivity anal- ysis to include uncertainties regarding osteological articulations in non-avian dinosaurs, including effects of inter-vertebral spacing and the orientation or ‘flare’ of the rib cage in MOR693. Results suggest body mass and inertial values are extremely uncertain and show a wide range in plausible values, whilst the CM (centre of mass) position is well constrained immediately in front and below the hip joint in MOR693, consistent with similar reconstructions of non-avian theropods. Karl T. -
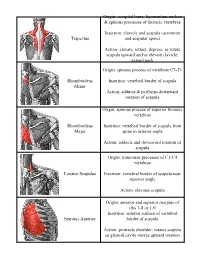
Trapezius Origin: Occipital Bone, Ligamentum Nuchae & Spinous Processes of Thoracic Vertebrae Insertion: Clavicle and Scapul
Origin: occipital bone, ligamentum nuchae & spinous processes of thoracic vertebrae Insertion: clavicle and scapula (acromion Trapezius and scapular spine) Action: elevate, retract, depress, or rotate scapula upward and/or elevate clavicle; extend neck Origin: spinous process of vertebrae C7-T1 Rhomboideus Insertion: vertebral border of scapula Minor Action: adducts & performs downward rotation of scapula Origin: spinous process of superior thoracic vertebrae Rhomboideus Insertion: vertebral border of scapula from Major spine to inferior angle Action: adducts and downward rotation of scapula Origin: transverse precesses of C1-C4 vertebrae Levator Scapulae Insertion: vertebral border of scapula near superior angle Action: elevates scapula Origin: anterior and superior margins of ribs 1-8 or 1-9 Insertion: anterior surface of vertebral Serratus Anterior border of scapula Action: protracts shoulder: rotates scapula so glenoid cavity moves upward rotation Origin: anterior surfaces and superior margins of ribs 3-5 Insertion: coracoid process of scapula Pectoralis Minor Action: depresses & protracts shoulder, rotates scapula (glenoid cavity rotates downward), elevates ribs Origin: supraspinous fossa of scapula Supraspinatus Insertion: greater tuberacle of humerus Action: abduction at the shoulder Origin: infraspinous fossa of scapula Infraspinatus Insertion: greater tubercle of humerus Action: lateral rotation at shoulder Origin: clavicle and scapula (acromion and adjacent scapular spine) Insertion: deltoid tuberosity of humerus Deltoid Action: -

The Pelvic and Hind Limb Anatomy of the Stem-Sauropodomorph Saturnalia Tupiniquim (Late Triassic, Brazil)
PaleoBios 23(2):1–30, July 15, 2003 © 2003 University of California Museum of Paleontology The pelvic and hind limb anatomy of the stem-sauropodomorph Saturnalia tupiniquim (Late Triassic, Brazil) MAX CARDOSO LANGER Department of Earth Sciences, University of Bristol, Wills Memorial Building, Queens Road, BS8 1RJ Bristol, UK. Current address: Departamento de Biologia, Universidade de São Paulo (USP), Av. Bandeirantes, 3900 14040-901 Ribeirão Preto, SP, Brazil; [email protected] Three partial skeletons allow a nearly complete description of the sacrum, pelvic girdle, and hind limb of the stem- sauropodomorph Saturnalia tupiniquim, from the Late Triassic Santa Maria Formation, South Brazil. The new morphological data gathered from these specimens considerably improves our knowledge of the anatomy of basal dinosaurs, providing the basis for a reassessment of various morphological transformations that occurred in the early evolution of these reptiles. These include an increase in the number of sacral vertebrae, the development of a brevis fossa, the perforation of the acetabulum, the inturning of the femoral head, as well as various modifications in the insertion of the iliofemoral musculature and the tibio-tarsal articulation. In addition, the reconstruction of the pelvic musculature of Saturnalia, along with a study of its locomotion pattern, indicates that the hind limb of early dinosaurs did not perform only a fore-and-aft stiff rotation in the parasagittal plane, but that lateral and medial movements of the leg were also present and important. INTRODUCTION sisting of most of the presacral vertebral series, both sides Saturnalia tupiniquim was described in a preliminary of the pectoral girdle, right humerus, partial right ulna, right fashion by Langer et al. -

The Nonavian Theropod Quadrate II: Systematic Usefulness, Major Trends and Cladistic and Phylogenetic Morphometrics Analyses
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/272162807 The nonavian theropod quadrate II: systematic usefulness, major trends and cladistic and phylogenetic morphometrics analyses Article · January 2014 DOI: 10.7287/peerj.preprints.380v2 CITATION READS 1 90 3 authors: Christophe Hendrickx Ricardo Araujo University of the Witwatersrand Technical University of Lisbon 37 PUBLICATIONS 210 CITATIONS 89 PUBLICATIONS 324 CITATIONS SEE PROFILE SEE PROFILE Octávio Mateus University NOVA of Lisbon 224 PUBLICATIONS 2,205 CITATIONS SEE PROFILE Some of the authors of this publication are also working on these related projects: Nature and Time on Earth - Project for a course and a book for virtual visits to past environments in learning programmes for university students (coordinators Edoardo Martinetto, Emanuel Tschopp, Robert A. Gastaldo) View project Ten Sleep Wyoming Jurassic dinosaurs View project All content following this page was uploaded by Octávio Mateus on 12 February 2015. The user has requested enhancement of the downloaded file. The nonavian theropod quadrate II: systematic usefulness, major trends and cladistic and phylogenetic morphometrics analyses Christophe Hendrickx1,2 1Universidade Nova de Lisboa, CICEGe, Departamento de Ciências da Terra, Faculdade de Ciências e Tecnologia, Quinta da Torre, 2829-516, Caparica, Portugal. 2 Museu da Lourinhã, 9 Rua João Luis de Moura, 2530-158, Lourinhã, Portugal. s t [email protected] n i r P e 2,3,4,5 r Ricardo Araújo P 2 Museu da Lourinhã, 9 Rua João Luis de Moura, 2530-158, Lourinhã, Portugal. 3 Huffington Department of Earth Sciences, Southern Methodist University, PO Box 750395, 75275-0395, Dallas, Texas, USA. -

Coracoid Process Anatomy: a Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B
Quantitative Assessment of the Coracoacromial and the Coracoclavicular Ligaments With 3-Dimensional Mapping of the Coracoid Process Anatomy: A Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B. Ferrari, M.D., George Sanchez, B.S., Alex W. Brady, M.Sc., Jonas Pogorzelski, M.D., M.H.B.A., George F. Lebus, M.D., Peter J. Millett, M.D., M.Sc., Robert F. LaPrade, M.D., Ph.D., and CAPT Matthew T. Provencher, M.D., M.C., U.S.N.R. Purpose: To perform a quantitative anatomic evaluation of the (1) coracoid process, specifically the attachment sites of the conjoint tendon, the pectoralis minor, the coracoacromial ligament (CAL), and the coracoclavicular (CC) ligaments in relation to pertinent osseous and soft tissue landmarks; (2) CC ligaments’ attachments on the clavicle; and (3) CAL attachment on the acromion in relation to surgically relevant anatomic landmarks to assist in planning of the Latarjet procedure, acromioclavicular (AC) joint reconstructions, and CAL resection distances avoiding iatrogenic injury to sur- rounding structures. Methods: Ten nonpaired fresh-frozen human cadaveric shoulders (mean age 52 years, range 33- 64 years) were included in this study. A 3-dimensional coordinate measuring device was used to quantify the location of pertinent bony landmarks and soft tissue attachment areas. The ligament and tendon attachment perimeters and center points on the coracoid, clavicle, and acromion were identified and subsequently dissected off the bone. Coordinates of points along the perimeters of attachment sites were used to calculate areas, whereas coordinates of center points were used to determine distances between surgically relevant attachment sites and pertinent bony landmarks.