Inheritance of Cross-Sensitivity in Sweet Corn to Herbicides Applied Postemergence
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2,4-Dichlorophenoxyacetic Acid
2,4-Dichlorophenoxyacetic acid 2,4-Dichlorophenoxyacetic acid IUPAC (2,4-dichlorophenoxy)acetic acid name 2,4-D Other hedonal names trinoxol Identifiers CAS [94-75-7] number SMILES OC(COC1=CC=C(Cl)C=C1Cl)=O ChemSpider 1441 ID Properties Molecular C H Cl O formula 8 6 2 3 Molar mass 221.04 g mol−1 Appearance white to yellow powder Melting point 140.5 °C (413.5 K) Boiling 160 °C (0.4 mm Hg) point Solubility in 900 mg/L (25 °C) water Related compounds Related 2,4,5-T, Dichlorprop compounds Except where noted otherwise, data are given for materials in their standard state (at 25 °C, 100 kPa) 2,4-Dichlorophenoxyacetic acid (2,4-D) is a common systemic herbicide used in the control of broadleaf weeds. It is the most widely used herbicide in the world, and the third most commonly used in North America.[1] 2,4-D is also an important synthetic auxin, often used in laboratories for plant research and as a supplement in plant cell culture media such as MS medium. History 2,4-D was developed during World War II by a British team at Rothamsted Experimental Station, under the leadership of Judah Hirsch Quastel, aiming to increase crop yields for a nation at war.[citation needed] When it was commercially released in 1946, it became the first successful selective herbicide and allowed for greatly enhanced weed control in wheat, maize (corn), rice, and similar cereal grass crop, because it only kills dicots, leaving behind monocots. Mechanism of herbicide action 2,4-D is a synthetic auxin, which is a class of plant growth regulators. -

WO 2012/125779 Al 20 September 2012 (20.09.2012) P O P C T
(12) INTERNATIONAL APPLICATION PUBLISHED UNDER THE PATENT COOPERATION TREATY (PCT) (19) World Intellectual Property Organization International Bureau (10) International Publication Number (43) International Publication Date WO 2012/125779 Al 20 September 2012 (20.09.2012) P O P C T (51) International Patent Classification: CA, CH, CL, CN, CO, CR, CU, CZ, DE, DK, DM, DO, A01N 43/90 (2006.01) DZ, EC, EE, EG, ES, FI, GB, GD, GE, GH, GM, GT, HN, HR, HU, ID, IL, IN, IS, JP, KE, KG, KM, KN, KP, KR, (21) International Application Number: KZ, LA, LC, LK, LR, LS, LT, LU, LY, MA, MD, ME, PCT/US20 12/029 153 MG, MK, MN, MW, MX, MY, MZ, NA, NG, NI, NO, NZ, (22) International Filing Date: OM, PE, PG, PH, PL, PT, QA, RO, RS, RU, RW, SC, SD, 15 March 2012 (15.03.2012) SE, SG, SK, SL, SM, ST, SV, SY, TH, TJ, TM, TN, TR, TT, TZ, UA, UG, US, UZ, VC, VN, ZA, ZM, ZW. (25) Filing Language: English (84) Designated States (unless otherwise indicated, for every (26) Publication Language: English kind of regional protection available): ARIPO (BW, GH, (30) Priority Data: GM, KE, LR, LS, MW, MZ, NA, RW, SD, SL, SZ, TZ, 61/453,202 16 March 201 1 (16.03.201 1) US UG, ZM, ZW), Eurasian (AM, AZ, BY, KG, KZ, MD, RU, TJ, TM), European (AL, AT, BE, BG, CH, CY, CZ, DE, (71) Applicant (for all designated States except US) : DOW DK, EE, ES, FI, FR, GB, GR, HR, HU, IE, IS, IT, LT, LU, AGROSCIENCES LLC [US/US]; 9330 Zionsville Road, LV, MC, MK, MT, NL, NO, PL, PT, RO, RS, SE, SI, SK, Indianapolis, IN 46268 (US). -

Solventless Formulation of Triclopyr Butoxyethyl
(19) & (11) EP 1 983 829 B1 (12) EUROPEAN PATENT SPECIFICATION (45) Date of publication and mention (51) Int Cl.: of the grant of the patent: A01N 43/40 (2006.01) A01N 25/30 (2006.01) 13.01.2010 Bulletin 2010/02 A01N 25/04 (2006.01) A01P 13/00 (2006.01) (21) Application number: 06837962.7 (86) International application number: PCT/US2006/044751 (22) Date of filing: 17.11.2006 (87) International publication number: WO 2007/094836 (23.08.2007 Gazette 2007/34) (54) SOLVENTLESS FORMULATION OF TRICLOPYR BUTOXYETHYL ESTER LÖSUNGSMITTELFREIE FORMULIERUNG VON TRICLOPYR-BUTOXYETHYLESTER FORMULE SANS SOLVANT D’ESTER DE BUTOXYETHYLE DU TRICYCLOPYR (84) Designated Contracting States: • POMPEO, Michael, P. DE ES FR GB IT Sumter, SC 29150 (US) (30) Priority: 15.02.2006 US 773417 P (74) Representative: Weiss, Wolfgang Weickmann & Weickmann (43) Date of publication of application: Patentanwälte 29.10.2008 Bulletin 2008/44 Postfach 86 08 20 81635 München (DE) (73) Proprietors: • Dow AgroSciences LLC (56) References cited: Indianapolis, WO-A-96/28027 WO-A-2004/093546 Indiana 46268-1054 (US) US-A1- 5 466 659 • Jensen, Jeffrey, Lee Brownsburg IN 46112 (US) • BOVEY RODNEY W ET AL: "Honey mesquite (Prosopis glandulosa) control by synergistic (72) Inventors: action of clopyralid: Triclopyr mixtures" WEED • JENSEN, Jeffrey, Lee SCIENCE, vol. 40, no. 4, 1992, pages 563-567, Brownsburg, IN 46112 (US) XP009080681 ISSN: 0043-1745 • KLINE, William, N., III • BEBAWI F F ET AL: "Impact of foliar herbicides Duluth, GA 30096 (US) on pod and seed behaviour of rust-infected • BURCH, Patrick, L. rubber vine (Cryptostegia grandiflora) plants" Christiansburg, VA 24073 (US) PLANT PROTECTION QUARTERLY, vol. -

China Releases New Maximum Residue Limits for Pesticides In
GB 2763-2016 THIS REPORT CONTAINS ASSESSMENTS OF COMMODITY AND TRADE ISSUES MADE BY USDA STAFF AND NOT NECESSARILY STATEMENTS OF OFFICIAL U.S. GOVERNMENT POLICY Voluntary - Public Date: 3/31/2017 GAIN Report Number: CH17016 China - Peoples Republic of Post: Beijing China Releases New Maximum Residue Limits for Pesticides in Food Report Categories: FAIRS Subject Report Approved By: Lisa Anderson Prepared By: FAS Staff Report Highlights: On December 18, 2016, the Chinese National Health and Family Planning Commission, Ministry of Agriculture, China Food and Drug Administration released the National Food Safety Standard - Maximum Residue Limits for Pesticides in Foods (GB 2763-2016). The standard will replace the current MRL Standard (GB 2763-2014) and will be implemented on June 18, 2017. This report provides an unofficial translation of the standard. Editors’ Note: The asterisk appearing in the MRL column means that the limit is a temporary MRL. A temporary MRL is usually set under the following four conditions: 1. The dietary risk assessment data is incomplete; 2. The Acceptable Daily Intake (ADI) is temporary (ADI is used as the basis for MRL setting); 3. There is no surveillance or analysis method for the MRL that complies with the standard requirements; 4. In emergency situations, the pesticide is approved to be used on un-registered crops. I GB 2763-2016 General Information: BEGIN TRANSLATION ICS 65.100 G 25 GB National Standard of the People’s Republic of China GB 2763—2016 Replacing GB 2763 - 2014 National food safety standard Maximum Residue Limits for Pesticides in Food General Information: National Health and Family Planning Commission Issued by: Ministry of Agriculture China Food and Drug Administration Issued on: 2016-12-18 Implementation:2017-06-18 II GB 2763-2016 Table of Content Preface ............................................................................................................................................................... -
Method Description
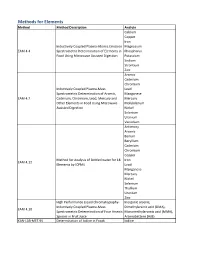
Methods for Elements Method Method Description Analyte Calcium Copper Iron Inductively Coupled Plasma-Atomic Emission Magnesium EAM 4.4 Spectrometric Determination of Elements in Phosphorus Food Using Microwave Assisted Digestion Potassium Sodium Strontium Zinc Arsenic Cadmium Chromium Inductively Coupled Plasma-Mass Lead Spectrometric Determination of Arsenic, Manganese EAM 4.7 Cadmium, Chromium, Lead, Mercury and Mercury Other Elements in Food Using Microwave Molybdenum Assisted Digestion Nickel Selenium Uranium Vanadium Antimony Arsenic Barium Beryllium Cadmium Chromium Copper Method for Analysis of Bottled water for 18 Iron EAM 4.12 Elements by ICPMS Lead Manganese Mercury Nickel Selenium Thallium Uranium Zinc High Performance Liquid Chromatography- Inorganic arsenic, Inductively Coupled Plasma-Mass Dimethylarsinic acid (DMA), EAM 4.10 Spectrometric Determination of Four Arsenic Monomethylarsonic acid (MMA), Species in Fruit Juice Arsenobetaine (AsB) KAN-LAB-MET.95 Determination of Iodine in Foods Iodine Methods for Radionuclides Method Method Description Analyte Determination of Strontium-90 in Foods by WEAC.RN.METHOD.2.0 Strontium-90 Internal Gas-Flow Proportional Counting Americium-241 Cesium-134 Cesium-137 Determination of Gamma-Ray Emitting Cobalt-60 WEAC.RN.METHOD.3.0 Radionuclides in Foods by High-Purity Potassium-40 Germanium Spectrometry Radium-226 Ruthenium-103 Ruthenium-106 Thorium-232 Methods for Pesticides/Industrial Chemicals Method Method Description Analyte Extraction Method: Analysis of Pesticides KAN-LAB-PES.53 and -

PPO-Inhibiting Herbicides and Structurally Relevant Schiff Bases: Evaluation of Inhibitory Activities Against Human Protoporphyrinogen Oxidase
processes Article PPO-Inhibiting Herbicides and Structurally Relevant Schiff Bases: Evaluation of Inhibitory Activities against Human Protoporphyrinogen Oxidase Milan Jakubek 1,2,3 , Michal Masaˇrík 1,2,4,5, Tomáš Bˇríza 1,2,3, Robert Kaplánek 1,2,3 , KateˇrinaVeselá 1,2,3, Nikita Abramenko 1 and Pavel Martásek 1,* 1 Department of Paediatrics and Inherited Metabolic Disorders, First Faculty of Medicine, Charles University and General University Hospital in Prague, 121 08 Prague, Czech Republic; [email protected] (M.J.); [email protected] (M.M.); [email protected] (T.B.); [email protected] (R.K.); [email protected] (K.V.); [email protected] (N.A.) 2 BIOCEV, First Faculty of Medicine, Charles University, 252 50 Vestec, Czech Republic 3 Department of Analytical Chemistry, Faculty of Chemical Engineering, University of Chemistry and Technology, 166 28 Prague, Czech Republic 4 Department of Physiology, Faculty of Medicine, Masaryk University, Kamenice 5, 625 00 Brno, Czech Republic 5 Department of Pathological Physiology, Faculty of Medicine, Masaryk University, Kamenice 5, 625 00 Brno, Czech Republic * Correspondence: [email protected] Abstract: The study of human protoporphyrinogen oxidase (hPPO) inhibition can contribute signif- icantly to a better understanding of some pathogeneses (e.g., porphyria, herbicide exposure) and the development of anticancer agents. Therefore, we prepared new potential inhibitors with Schiff Citation: Jakubek, M.; Masaˇrík,M.; base structural motifs (2-hydroxybenzaldehyde-based Schiff bases 9–13 and chromanone derivatives Bˇríza,T.; Kaplánek, R.; Veselá, K.; 17–19) as structurally relevant to PPO herbicides. The inhibitory activities (represented by the half Abramenko, N.; Martásek, P. -

(Bio)Sensors for Pesticides Detection Using Screen-Printed Electrodes
biosensors Review Electrochemical (Bio)Sensors for Pesticides Detection Using Screen-Printed Electrodes Beatriz Pérez-Fernández , Agustín Costa-García y and Alfredo de la Escosura- Muñiz * NanoBioAnalysis Group-Department of Physical and Analytical Chemistry, University of Oviedo, Julián Clavería 8, 33006 Oviedo, Spain * Correspondence: [email protected]; Tel.: +34-985-103-521 In Memoriam to Prof. Agustín Costa-García. y Received: 12 March 2020; Accepted: 30 March 2020; Published: 2 April 2020 Abstract: Pesticides are among the most important contaminants in food, leading to important global health problems. While conventional techniques such as high-performance liquid chromatography (HPLC) and mass spectrometry (MS) have traditionally been utilized for the detection of such food contaminants, they are relatively expensive, time-consuming and labor intensive, limiting their use for point-of-care (POC) applications. Electrochemical (bio)sensors are emerging devices meeting such expectations, since they represent reliable, simple, cheap, portable, selective and easy to use analytical tools that can be used outside the laboratories by non-specialized personnel. Screen-printed electrodes (SPEs) stand out from the variety of transducers used in electrochemical (bio)sensing because of their small size, high integration, low cost and ability to measure in few microliters of sample. In this context, in this review article, we summarize and discuss about the use of SPEs as analytical tools in the development of (bio)sensors for pesticides of interest for food control. Finally, aspects related to the analytical performance of the developed (bio)sensors together with prospects for future improvements are discussed. Keywords: screen-printed electrodes; electrochemical (bio)sensors; pesticides; point-of-care; food control 1. -

Herbicide Resistance: Toward an Understanding of Resistance Development and the Impact of Herbicide-Resistant Crops William K
Weed Science 2012 Special Issue:2–30 Herbicide Resistance: Toward an Understanding of Resistance Development and the Impact of Herbicide-Resistant Crops William K. Vencill, Robert L. Nichols, Theodore M. Webster, John K. Soteres, Carol Mallory-Smith, Nilda R. Burgos, William G. Johnson, and Marilyn R. McClelland* Table of Contents and how they affect crop production and are affected by management practices, and to present the environmental impacts Executive Summary……………………………………… 2 of herbicide-resistant crops. This paper will summarize aspects of I. Introduction: A Summary of Weed Science Practices herbicide resistance in five different sections: (1) a description of and Concepts………………………………………… 3 basic weed science management practices and concepts, (2) II. Resistance and Tolerance in Weed Science………… 12 definitions of resistance and tolerance in weed science, (3) envi- III. Environmental Impacts of Herbicide Resistance in ronmental impacts of herbicide-resistant crops, (4) strategies for Crops………………………………………………… 15 management of weed species shifts and herbicide-resistant weeds IV. Strategies for Managing Weed Species Shifts and Devel- and adoption by the agricultural community, and (5) gene-flow opment of Herbicide-Resistant Weeds…………………… 16 potential from herbicide-resistant crops. V. Gene Flow from Herbicide-Resistant Crops………… 19 Literature Cited…………………………………………… 24 Section 1: Introduction. To avoid or delay the development of resistant weeds, a diverse, integrated program of weed management practices is required to minimize reliance -

Metabolism of 2,4-Dichlorophenoxyacetic Acid Contributes to Resistance in a Common Waterhemp (Amaranthus Tuberculatus) Population
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Agronomy & Horticulture -- Faculty Publications Agronomy and Horticulture Department 2017 Metabolism of 2,4-dichlorophenoxyacetic acid contributes to resistance in a common waterhemp (Amaranthus tuberculatus) population Marcelo R. A. Figueiredo Colorado State University Lacy J. Leibhart University of Nebraska-Lincoln Zachary J. Reicher University of Nebraska-Lincoln Patrick J. Tranel University of Illinois Scott J. Nissen Colorado State University Follow this and additional works at: https://digitalcommons.unl.edu/agronomyfacpub See next page for additional authors Part of the Agricultural Science Commons, Agriculture Commons, Agronomy and Crop Sciences Commons, Botany Commons, Horticulture Commons, Other Plant Sciences Commons, and the Plant Biology Commons Figueiredo, Marcelo R. A.; Leibhart, Lacy J.; Reicher, Zachary J.; Tranel, Patrick J.; Nissen, Scott J.; Westra, Philip; Bernards, Mark L.; Kruger, Greg R.; Gaines, Todd A.; and Jugulam, Mithila, "Metabolism of 2,4-dichlorophenoxyacetic acid contributes to resistance in a common waterhemp (Amaranthus tuberculatus) population" (2017). Agronomy & Horticulture -- Faculty Publications. 1057. https://digitalcommons.unl.edu/agronomyfacpub/1057 This Article is brought to you for free and open access by the Agronomy and Horticulture Department at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Agronomy & Horticulture -- Faculty Publications by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Authors Marcelo R. A. Figueiredo, Lacy J. Leibhart, Zachary J. Reicher, Patrick J. Tranel, Scott J. Nissen, Philip Westra, Mark L. Bernards, Greg R. Kruger, Todd A. Gaines, and Mithila Jugulam This article is available at DigitalCommons@University of Nebraska - Lincoln: https://digitalcommons.unl.edu/ agronomyfacpub/1057 Metabolism of 2,4-dichlorophenoxyacetic acid contributes to resistance in a common waterhemp (Amaranthus tuberculatus) population Marcelo R. -

Alternative Strategies for Weed Control in Creeping Bentgrass
University of Tennessee, Knoxville TRACE: Tennessee Research and Creative Exchange Doctoral Dissertations Graduate School 8-2014 Alternative Strategies for Weed Control in Creeping Bentgrass Matthew Thomas Elmore University of Tennessee - Knoxville, [email protected] Follow this and additional works at: https://trace.tennessee.edu/utk_graddiss Part of the Weed Science Commons Recommended Citation Elmore, Matthew Thomas, "Alternative Strategies for Weed Control in Creeping Bentgrass. " PhD diss., University of Tennessee, 2014. https://trace.tennessee.edu/utk_graddiss/2820 This Dissertation is brought to you for free and open access by the Graduate School at TRACE: Tennessee Research and Creative Exchange. It has been accepted for inclusion in Doctoral Dissertations by an authorized administrator of TRACE: Tennessee Research and Creative Exchange. For more information, please contact [email protected]. To the Graduate Council: I am submitting herewith a dissertation written by Matthew Thomas Elmore entitled "Alternative Strategies for Weed Control in Creeping Bentgrass." I have examined the final electronic copy of this dissertation for form and content and recommend that it be accepted in partial fulfillment of the requirements for the degree of Doctor of Philosophy, with a major in Plants, Soils, and Insects. James T. Brosnan, Major Professor We have read this dissertation and recommend its acceptance: John C. Sorochan, Dean A. Kopsell, Thomas C. Mueller, Michael D. Best Accepted for the Council: Carolyn R. Hodges Vice Provost and Dean of the Graduate School (Original signatures are on file with official studentecor r ds.) Alternative Strategies for Weed Control in Creeping Bentgrass A Dissertation Presented for the Doctor of Philosophy Degree The University of Tennessee, Knoxville Matthew Thomas Elmore August 2014 Copyright © 2014 by Matthew Elmore All rights reserved ii ACKNOWLEDGEMENTS For their support and guidance throughout my time at the University of Tennessee, I thank my committee members: Dr. -

Physiological, Biochemical and Molecular Characterization of Multiple Herbicide Resistance in Palmer Amaranth (Amaranthus Palmeri)
PHYSIOLOGICAL, BIOCHEMICAL AND MOLECULAR CHARACTERIZATION OF MULTIPLE HERBICIDE RESISTANCE IN PALMER AMARANTH (AMARANTHUS PALMERI) by SRIDEVI NAKKA M.S., Acharya N.G. Ranaga Agricultural University, 2005 M.S., University of Illinois, 2008 AN ABSTRACT OF A DISSERTATION submitted in partial fulfillment of the requirements for the degree DOCTOR OF PHILOSOPHY Department of Agronomy College of Agriculture KANSAS STATE UNIVERSITY Manhattan, Kansas 2016 Abstract Palmer amaranth (Amaranthus palmeri) is one of the most aggressive, troublesome and damaging broadleaf weeds in many cropping systems including corn, soybean, cotton, and grain sorghum causing huge yield losses across the USA. As a result of extensive and intensive selection of pre- and -post emergence herbicides, Palmer amaranth has evolved resistance to multiple herbicide modes of action, microtubule-, 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS)-, acetolactate synthase (ALS)-, photosystem II (PS II)-, hydroxyphenylpyruvate dioxygenase (HPPD)- and more recently to protoporphyrinogen oxidase (PPO)-inhibitors. A Palmer amaranth population from Kansas was found resistant to HPPD-, PS II-, and ALS- inhibitors. The overall objective of this research was to investigate the target-site and/or non- target-site resistance mechanisms in Palmer amaranth from KS (KSR) to mesotrione (HPPD- inhibitor), atrazine (PS II-inhibitor), and chlorsulfuron (ALS-inhibitor) relative to known susceptible Palmer amaranth from Mississippi (MSS) and KS (KSS). Whole plant dose-response assays showed high level of resistance in KSR to mesotrione, atrazine and chlorsulfuron. KSR was 10-18, 178-237 and >275 fold more resistant to mesotrione, atrazine, and chlorsulfuron, respectively, compared to MSS and KSS. Metabolism studies using [14C] labeled mesotrione and atrazine demonstrated non-target-site resistance to both herbicides, particularly, enhanced metabolism of [14C] mesotrione likely mediated by cytochrome P450 monooxygenases and rapid degradation of [14C] atrazine by glutathione S-transferases (GSTs). -

Studies on the Varietal Susceptibility in Winter
STUDIES ON THE VARIETAL SUSCEPTIBILITY IN WINTER WHEAT TO SUBSTITUTED PHENYLUREA HERBICIDES A Thesis submitted by REZA EMAMI-SARAVI a candidate for the degree of Doctor of Philosophy in BIOCHEMISTRY Department of Biochemistry Royal Holloway College University of London Egham Hill Egham January 1979 Surrey ProQuest Number: 10097467 All rights reserved INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed, a note will indicate the deletion. uest. ProQuest 10097467 Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author. All rights reserved. This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC. ProQuest LLC 789 East Eisenhower Parkway P.O. Box 1346 Ann Arbor, Ml 48106-1346 (i) ACKN OWLEDGEMENTS My sincere thanks and gratitude must go to my supervisor. Dr. W.J. Owen, for introducing me to such an interesting topic, for his help, encouragement, and patience throughout my research, and for his guidance in the preparation of this thesis. Secondly, I wish to thank Professor J.B. Pridham and Dr. W.A. Stevens for their interest, advice and helpful discussions throughout the duration of this work. My gratitude is also extended to all other members of staff and to students of this College who have helped me either practically or theoretically during my postgraduate research and in the preparation of this thesis.