Activates the Lectin Complement Pathway Human M-Ficolin Is A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Membrane Complement Regulatory Protein CD59 and Its Association with Rheumatoid Arthritis and Systemic Lupus Erythematosus
Current Medicine Research and Practice 9 (2019) 182e188 Contents lists available at ScienceDirect Current Medicine Research and Practice journal homepage: www.elsevier.com/locate/cmrp Review Article The membrane complement regulatory protein CD59 and its association with rheumatoid arthritis and systemic lupus erythematosus * Nibhriti Das a, Devyani Anand a, Bintili Biswas b, Deepa Kumari c, Monika Gandhi c, a Department of Biochemistry, All India Institute of Medical Sciences, New Delhi 110029, India b Department of Zoology, Ramjas College, University of Delhi, India c University School of Biotechnology, Guru Gobind Singh Indraprastha University, India article info abstract Article history: The complement cascade consisting of about 50 soluble and cell surface proteins is activated in auto- Received 8 May 2019 immune inflammatory disorders. This contributes to the pathological manifestations in these diseases. In Accepted 30 July 2019 normal health, the soluble and membrane complement regulatory proteins protect the host against Available online 5 August 2019 complement-mediated self-tissue injury by controlling the extent of complement activation within the desired limits for the host's benefit. CD59 is a membrane complement regulatory protein that inhibits the Keywords: formation of the terminal complement complex or membrane attack complex (C5b6789n) which is CD59 generated on complement activation by any of the three pathways, namely, the classical, alternative, and RA SLE the mannose-binding lectin pathway. Animal experiments and human studies have suggested impor- Pathophysiology tance of membrane complement proteins including CD59 in the pathophysiology of rheumatoid arthritis Disease marker (RA) and systemic lupus erythematosus (SLE). Here is a brief review on CD59 and its distribution, structure, functions, and association with RA and SLE starting with a brief introduction on the com- plement system, its activation, the biological functions, and relations of membrane complement regu- latory proteins, especially CD59, with RA and SLE. -

Complement Recognition Pathways in Renal Transplantation
BRIEF REVIEW www.jasn.org Complement Recognition Pathways in Renal Transplantation Christopher L. Nauser, Conrad A. Farrar, and Steven H. Sacks Medical Research Council Centre for Transplantation, Division of Transplantation Immunology and Mucosal Biology, King’s College London, National Health Service Guy’s and St. Thomas’ Trust, London, United Kingdom ABSTRACT The complement system, consisting of soluble and cell membrane–bound compo- weigh its effect on organ injury and nents of the innate immune system, has defined roles in the pathophysiology of renal rejection with dampening the antimi- allograft rejection. Notably, the unavoidable ischemia-reperfusion injury inherent to crobial functions of the complement sys- transplantation is mediated through the terminal complement activation products tem, such as opsonisation, cell lysis, and C5a and C5b-9. Furthermore, biologically active fragments C3a and C5a, produced recruitment of neutrophils and other in- during complement activation, can modulate both antigen presentation and T cell flammatory cells.7 It is our belief that priming, ultimately leading to allograft rejection. Earlier work identified renal tubule therapeutic strategies can be designed cell synthesis of C3, rather than hepatic synthesis of C3, as the primary source of C3 to specificallytargetthekeyinitiators driving these effects. Recent efforts have focused on identifying the local triggers of of complement activation at the relevant complement activation. Collectin-11, a soluble C-type lectin expressed in renal tis- location, such as complement-binding sue, has been implicated as an important trigger of complement activation in renal anti-HLA antibodies in the vascular tissue. In particular, collectin-11 has been shown to engage L-fucose at sites of compartment or the initiator(s) of local ischemic stress, activating the lectin complement pathway and directing the innate complement activation in the extravas- immune response to the distressed renal tubule. -

Activation Regulates Alternative Pathway Complement Necrotic
Properdin Binds to Late Apoptotic and Necrotic Cells Independently of C3b and Regulates Alternative Pathway Complement Activation This information is current as of September 27, 2021. Wei Xu, Stefan P. Berger, Leendert A. Trouw, Hetty C. de Boer, Nicole Schlagwein, Chantal Mutsaers, Mohamed R. Daha and Cees van Kooten J Immunol 2008; 180:7613-7621; ; doi: 10.4049/jimmunol.180.11.7613 Downloaded from http://www.jimmunol.org/content/180/11/7613 References This article cites 56 articles, 27 of which you can access for free at: http://www.jimmunol.org/ http://www.jimmunol.org/content/180/11/7613.full#ref-list-1 Why The JI? Submit online. • Rapid Reviews! 30 days* from submission to initial decision • No Triage! Every submission reviewed by practicing scientists by guest on September 27, 2021 • Fast Publication! 4 weeks from acceptance to publication *average Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2008 by The American Association of Immunologists All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. The Journal of Immunology Properdin Binds to Late Apoptotic and Necrotic Cells Independently of C3b and Regulates Alternative Pathway Complement Activation1,2 Wei Xu,* Stefan P. Berger,* Leendert A. -

An Anticomplement Agent That Homes to the Damaged Brain and Promotes Recovery After Traumatic Brain Injury in Mice
An anticomplement agent that homes to the damaged brain and promotes recovery after traumatic brain injury in mice Marieta M. Rusevaa,1,2, Valeria Ramagliab,1, B. Paul Morgana, and Claire L. Harrisa,3 aInstitute of Infection and Immunity, School of Medicine, Cardiff University, Cardiff CF14 4XN, United Kingdom; and bDepartment of Genome Analysis, Academic Medical Center, Amsterdam 1105 AZ, The Netherlands Edited by Douglas T. Fearon, Cornell University, Cambridge, United Kingdom, and approved September 29, 2015 (received for review July 15, 2015) Activation of complement is a key determinant of neuropathology to rapidly and specifically inhibit MAC at sites of complement and disability after traumatic brain injury (TBI), and inhibition is activation, and test its therapeutic potential in experimental TBI. neuroprotective. However, systemic complement is essential to The construct, termed CD59-2a-CRIg, comprises CD59a linked fight infections, a critical complication of TBI. We describe a to CRIg via the murine IgG2a hinge. CD59a prevents assembly targeted complement inhibitor, comprising complement receptor of MAC in cell membranes (16), whereas CRIg binds C3b/iC3b of the Ig superfamily (CRIg) fused with complement regulator CD59a, deposited at sites of complement activation (17). The IgG2a designed to inhibit membrane attack complex (MAC) assembly at hinge promotes dimerization to increase ligand avidity. CD59- sites of C3b/iC3b deposition. CRIg and CD59a were linked via the 2a-CRIg protected in the TBI model, demonstrating that site- IgG2a hinge, yielding CD59-2a-CRIg dimer with increased iC3b/C3b targeted anti-MAC therapeutics may be effective in prevention binding avidity and MAC inhibitory activity. CD59-2a-CRIg inhibited of secondary neuropathology and improve neurologic recovery MAC formation and prevented complement-mediated lysis in vitro. -

Supplementary Table 1: Adhesion Genes Data Set
Supplementary Table 1: Adhesion genes data set PROBE Entrez Gene ID Celera Gene ID Gene_Symbol Gene_Name 160832 1 hCG201364.3 A1BG alpha-1-B glycoprotein 223658 1 hCG201364.3 A1BG alpha-1-B glycoprotein 212988 102 hCG40040.3 ADAM10 ADAM metallopeptidase domain 10 133411 4185 hCG28232.2 ADAM11 ADAM metallopeptidase domain 11 110695 8038 hCG40937.4 ADAM12 ADAM metallopeptidase domain 12 (meltrin alpha) 195222 8038 hCG40937.4 ADAM12 ADAM metallopeptidase domain 12 (meltrin alpha) 165344 8751 hCG20021.3 ADAM15 ADAM metallopeptidase domain 15 (metargidin) 189065 6868 null ADAM17 ADAM metallopeptidase domain 17 (tumor necrosis factor, alpha, converting enzyme) 108119 8728 hCG15398.4 ADAM19 ADAM metallopeptidase domain 19 (meltrin beta) 117763 8748 hCG20675.3 ADAM20 ADAM metallopeptidase domain 20 126448 8747 hCG1785634.2 ADAM21 ADAM metallopeptidase domain 21 208981 8747 hCG1785634.2|hCG2042897 ADAM21 ADAM metallopeptidase domain 21 180903 53616 hCG17212.4 ADAM22 ADAM metallopeptidase domain 22 177272 8745 hCG1811623.1 ADAM23 ADAM metallopeptidase domain 23 102384 10863 hCG1818505.1 ADAM28 ADAM metallopeptidase domain 28 119968 11086 hCG1786734.2 ADAM29 ADAM metallopeptidase domain 29 205542 11085 hCG1997196.1 ADAM30 ADAM metallopeptidase domain 30 148417 80332 hCG39255.4 ADAM33 ADAM metallopeptidase domain 33 140492 8756 hCG1789002.2 ADAM7 ADAM metallopeptidase domain 7 122603 101 hCG1816947.1 ADAM8 ADAM metallopeptidase domain 8 183965 8754 hCG1996391 ADAM9 ADAM metallopeptidase domain 9 (meltrin gamma) 129974 27299 hCG15447.3 ADAMDEC1 ADAM-like, -

Complement Inactivation Strategy of Staphylococcus Aureus Using Decay-Accelerating Factor and the Response of Infected Hacat Cells
International Journal of Molecular Sciences Article Complement Inactivation Strategy of Staphylococcus aureus Using Decay-Accelerating Factor and the Response of Infected HaCaT Cells Kyoung Ok Jang 1, Youn Woo Lee 1, Hangeun Kim 2,* and Dae Kyun Chung 1,2,3,* 1 Graduate School of Biotechnology, Kyung Hee University, Yongin 17104, Korea; [email protected] (K.O.J.); [email protected] (Y.W.L.) 2 Research and Development Center, Skin Biotechnology Center Inc., Yongin 17104, Korea 3 Skin Biotechnology Center, Kyung Hee University, Suwon 16229, Korea * Correspondence: [email protected] (H.K.); [email protected] (D.K.C.); Tel.: +82-31-201-2487 (H.K.); +82-31-888-6170 (D.K.C.) Abstract: Staphylococcus aureus is a species of Gram-positive staphylococcus. It can cause sinusitis, respiratory infections, skin infections, and food poisoning. Recently, it was discovered that S. aureus infects epithelial cells, but the interaction between S. aureus and the host is not well known. In this study, we confirmed S. aureus to be internalized by HaCaT cells using the ESAT-6-like protein EsxB and amplified within the host over time by escaping host immunity. S. aureus increases the expression of decay-accelerating factor (CD55) on the surfaces of host cells, which inhibits the activation of the complement system. This mechanism makes it possible for S. aureus to survive in host cells. S. aureus, sufficiently amplified within the host, is released through the initiation of cell death. On the other hand, the infected host cells increase their surface expression of UL16 binding protein 1 to inform Citation: Jang, K.O.; Lee, Y.W.; Kim, immune cells that they are infected and try to be eliminated. -

Recognition of Microbial Glycans by Soluble Human Lectins
Available online at www.sciencedirect.com ScienceDirect Recognition of microbial glycans by soluble human lectins 3 1 1,2 Darryl A Wesener , Amanda Dugan and Laura L Kiessling Human innate immune lectins that recognize microbial glycans implicated in the regulation of microbial colonization and can conduct microbial surveillance and thereby help prevent in protection against infection. Seminal research on the infection. Structural analysis of soluble lectins has provided acute response to bacterial infection led to the identifica- invaluable insight into how these proteins recognize their tion of secreted factors that include C-reactive protein cognate carbohydrate ligands and how this recognition gives (CRP) and mannose-binding lectin (MBL) [1,3]. Both rise to biological function. In this opinion, we cover the CRP and MBL can recognize carbohydrate antigens on structural features of lectins that allow them to mediate the surface of pathogens, including Streptococcus pneumo- microbial recognition, highlighting examples from the collectin, niae and Staphylococcus aureus and then promote comple- Reg protein, galectin, pentraxin, ficolin and intelectin families. ment-mediated opsonization and cell killing [4]. Since These analyses reveal how some lectins (e.g., human intelectin- these initial observations, other lectins have been impli- 1) can recognize glycan epitopes that are remarkably diverse, cated in microbial recognition. Like MBL some of these yet still differentiate between mammalian and microbial proteins are C-type lectins, while others are members of glycans. We additionally discuss strategies to identify lectins the ficolin, pentraxin, galectin, or intelectin families. that recognize microbial glycans and highlight tools that Many of the lectins that function in microbial surveillance facilitate these discovery efforts. -

Complement Herbert L
Host Defense 2011 Complement Herbert L. Mathews, Ph.D. COMPLEMENT Date: 4/11/11 Reading Assignment: Janeway’s Immunobiology, 7th Edition, pp. 54-55, 61-82, 406- 409, 514-515. Figures: (Unless otherwise noted) Janeway’s Immunobiology, 7th Edition, Murphy et al., Garland Publishing. KEY CONCEPTS AND LEARNING OBJECTIVES You will be able to describe the mechanism and consequences of the activation of the complement system. To attain the goals for these lectures you will be able to: a. List the components of the complement system. b. Describe the three activation pathways for complement. c. Explain the consequences of complement activation. d. Describe the consequence of complement deficiency. Page 1 Host Defense 2011 Complement Herbert L. Mathews, Ph.D. CONTENT SUMMARY Introduction Nomenclature Activation of Complement The classical pathway The mannan-binding lectin pathway The alternative pathway Biological Consequence of Complement Activation Cell lysis and viral neutralization Opsonization Clearance of Immune Complexes Inflammation Regulation of Complement Activation Human Complement Component Deficiencies Page 2 Host Defense 2011 Complement Herbert L. Mathews, Ph.D. Introduction The complement system is a group of more than 30 plasma and membrane proteins that play a critical role in host defense. When activated, complement components interact in a highly regulated fashion to generate products that: Recruit inflammatory cells (promoting inflammation). Opsonize microbial pathogens and immune complexes (facilitating antigen clearance). Kill microbial pathogens (via a lytic mechanism known as the membrane attack complex). Generate an inflammatory response. Complement activation takes place on antigenic surfaces. However, the activation of complement generates several soluble fragments that have important biologic activity. -
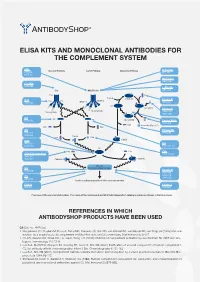
Elisa Kits and Monoclonal Antibodies for the Complement System
ELISA KITS AND MONOCLONAL ANTIBODIES FOR THE COMPLEMENT SYSTEM MBL ClassicalÊPathway LectinÊPathway AlternativeÊPathway H-Ficolin KITÊ029 RIGÊ334 HYBÊ131 M-Ficolin ABSÊ036 C1-INH HYBÊ288 L-Ficolin C1q MBL/Ficolin ABSÊ005 C1q B H2O C3Ê(H2O) FactorÊB C1s C1s C1r MASP-2 D ABSÊ002 MASP-1 HAVÊ005 C3 C3Ê(H O)ÊB 2 FactorÊD Carbohydrate Carbohydrate GAUÊ010 GAUÊ008 C2 C2 C4 C3Ê(H O)ÊBb HYBÊ050 2 Ba FactorÊBa/b HYBÊ008 C3a ProperdinÊ(FactorÊP) C2b C4a C3 Properdin HAVÊ004 HYBÊ0393 HYBÊ005 C4b2a C2a C4b C3bBb C3d HAVÊ003 C4 HYBÊ030 HYBÊ162-04 C3 C3a C3a C4b C3a/C3aÊ(desArg) HYBÊ162-02 GAUÊ013 GAUÊ017 C4b2a3b C3bBb3b C5Ê-ÊC9 C5 MembraneÊattackÊcomplex HYBÊ029 FactorÊH HYBÊ268 GAUÊ018 C9 GAUÊ020 ABSÊ004 LysisÊorÊphagocytosisÊofÊtheÊmicroorganism C5b-9 FactorÊI DIAÊ011 HYBÊ061 OverviewÊofÊtheÊcomplementÊsystem.ÊTheÊnameÊofÊtheÊcomponentÊandÊBioPortoÊDiagnostics'ÊcatalogÊnumberÊareÊshownÊinÊtheÊblueÊboxes. REFERENCES IN WHICH ANTIBODYSHOP PRODUCTS HAVE BEEN USED C2 Cat. no. HYB 050 Rooijakkers SH, Ruyken M, Roos A, Daha MR, Presanis JS, Sim RB, van Wamel WJ, van Kessel KP, van Strijp JA (2005) Immune evasion by a staphylococcal complement inhibitor that acts on C3 convertases. Nat Immunol 6:920-7. Oh KS, Kweon MH, Rhee KH, Ho Lee K, Sung HC (2003) Inhibition of complement activation by recombinant Sh-CRIT-ed1 ana- logues. Immunology 110:73-9. Laich A, Moffatt B, Wong KHN, Hickling TP, Koch C, Sim RB (2001) Purifi cation of second component of human complement, C2, by antibody affi nity chromatography. Intern J Bio-Chromatography 6:151-162. Laich A, Sim RB (2001) Complement C4bC2 complex formation: an investigation by surface plasmon resonance. Biochim Bio- phys Acta 1544:96-112. Stenbaek EI, Koch C, Barkholt V, Welinder KG (1986) Human complement component C2: production and characterization of polyclonal and monoclonal antibodies against C2. -

Table 1. Swine Proteins Identified As Differentially Expressed at 24Dpi in OURT 88/3 Infected Animals
Table 1. swine proteins identified as differentially expressed at 24dpi in OURT 88/3 infected animals. Gene name Protein ID Protein Name -Log p-value control vs A_24DPI Difference control Vs A_24DPI F8 K7GL28 Coagulation factor VIII 2.123919902 5.42533493 PPBP F1RUL6 C-X-C motif chemokine 3.219079808 4.493174871 SDPR I3LDR9 Caveolae associated protein 2 2.191007299 4.085711161 IGHG L8B0X5 IgG heavy chain 2.084611488 -4.282530149 LOC100517145 F1S3H9 Complement C3 (LOC100517145) 3.885740476 -4.364484406 GOLM1 F1S4I1 Golgi membrane protein 1 1.746130664 -4.767168681 FCN2 I3L5W3 Ficolin-2 2.937884686 -6.029483795 Table 2. swine proteins identified as differentially expressed at 7dpi in Benin ΔMGF infected animals. Gene name Protein ID Protein Name -Log p-value control vs B_7DPI Difference control Vs B_7DPI A0A075B7I5 Ig-like domain-containing protein 1.765578164 -3.480728149 ATP5A1 F1RPS8_PIG ATP synthase subunit alpha 2.270386995 3.270935059 LOC100627396 F1RX35_PIG Fibrinogen C-terminal domain-containing protein 2.211242648 3.967363358 LOC100514666;LOC102158263 F1RX36_PIG Fibrinogen alpha chain 2.337934993 3.758180618 FGB F1RX37_PIG Fibrinogen beta chain 2.411948004 4.03753376 PSMA8 F1SBA5_PIG Proteasome subunit alpha type 1.473601007 -3.815182686 ACAN F1SKR0_PIG Aggrecan core protein 1.974489764 -3.726634026 TFG F1SL01_PIG PB1 domain-containing protein 1.809215274 -3.131304741 LOC100154408 F1SSL6_PIG Proteasome subunit alpha type 1.701949053 -3.944885254 PSMA4 F2Z528_PIG Proteasome subunit alpha type 2.045768185 -4.502977371 PSMA5 F2Z5K2_PIG -

Targeting of Mannan-Binding Lectin-Associated Serine Protease-2 Confers Protection from Myocardial and Gastrointestinal Ischemia/Reperfusion Injury
Targeting of mannan-binding lectin-associated serine protease-2 confers protection from myocardial and gastrointestinal ischemia/reperfusion injury Wilhelm J. Schwaeblea,1, Nicholas J. Lyncha, James E. Clarkb, Michael Marberb, Nilesh J. Samanic, Youssif Mohammed Alia,d, Thomas Dudlere, Brian Parente, Karl Lhottaf, Russell Wallisa, Conrad A. Farrarg, Steven Sacksg, Haekyung Leeh, Ming Zhangh, Daisuke Iwakii, Minoru Takahashii, Teizo Fujitai, Clark E. Tedforde, and Cordula M. Stovera Departments of aInfection, Immunity, and Inflammation and cCardiovascular Sciences, University of Leicester, Leicester LE1 9HN, United Kingdom; bBritish Heart Foundation Centre and gMedical Research Council Centre for Transplantation and National Institute for Health Research Biomedical Research Centre at Guy’s and St. Thomas’ National Health Service Foundation Trust, King’s College London, London SE1 9RT, United Kingdom; dFaculty of Pharmacy, Department of Microbiology, University of Mansoura, Mansoura 35516, Egypt; eOmeros Corporation, Seattle, WA 98101; fLandeskrankenhaus Feldkirch, 6807 Feldkirch, Austria; hDepartment of Anesthesiology, State University of New York-Downstate Medical Center, New York, NY 11203; and iDepartment of Immunology, Fukushima Medical University, Fukushima 960-1295, Japan Edited* by Douglas T. Fearon, University of Cambridge School of Clinical Medicine, Cambridge, United Kingdom, and approved March 16, 2011 (received for review February 1, 2011) Complement research experienced a renaissance with the discovery aberrant glycosylation -

Regenerative Therapy 6 (2017) 52E64
Regenerative Therapy 6 (2017) 52e64 Contents lists available at ScienceDirect Regenerative Therapy journal homepage: http://www.elsevier.com/locate/reth Original Article Phase I clinical study of liver regenerative therapy for cirrhosis by intrahepatic arterial infusion of freshly isolated autologous adipose tissue-derived stromal/stem (regenerative) cell Yoshio Sakai a, d, Masayuki Takamura b, c, Akihiro Seki c, Hajime Sunagozaka a, c, Takeshi Terashima a, Takuya Komura c, Masatoshi Yamato a, c, Masaki Miyazawa c, Kazunori Kawaguchi a, Alessandro Nasti c, Hatsune Mochida c, Soichiro Usui b, c, * Nobuhisa Otani c, Takahiro Ochiya e, Takashi Wada d, Masao Honda a, Shuichi Kaneko a, c, a Department of Gastroenterology, Kanazawa University Hospital, Japan b Department of Cardiology, Kanazawa University Hospital, Japan c System Biology, Kanazawa University, Japan d Department of Laboratory Medicine, Kanazawa University, Japan e Division of Molecular and Cellular Medicine, National Cancer Institute, Japan article info abstract Article history: Introduction: Adipose tissue stromal cells contain a substantial number of mesenchymal stem cells. As Received 14 September 2016 such, their application to regeneration of miscellaneous impaired organs has attracted much attention. Received in revised form Methods: We designed a clinical study to investigate freshly isolated autologous adipose tissue-derived 12 November 2016 stromal/stem (regenerative) cell (ADRC) therapy for liver cirrhosis and conducted treatment in four Accepted 8 December 2016 cirrhotic patients. ADRCs were isolated from autologous subcutaneous adipose tissue obtained by the liposuction method, followed with use of the Celution system adipose tissue dissociation device. The Keywords: primary endpoint is assessment of safety one month after treatment. We also characterized the obtained Adipose tissue-derived regenerative cells Stromal cells ADRCs.