Temperature: Cold
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

1. TAKAKIACEAE S. Hattori & Inoue
1. TAKAKIACEAE S. Hattori & Inoue John R. Spence Wilfred B. Schofield Stems erect, arising sympodially from creeping pale or white stolons bearing clusters of beaked slime cells, rhizoids absent, stem in cross section differentiated into outer layer of smaller, thick- walled cells and cortex of larger thin-walled cells, sometimes with small central cells, these often with trigones, outer cells with simple oil droplets. Leaves typically 3-ranked, terete, forked, of (1–)2–4 terete segments, segments sometimes connate at base, in cross section of 3–5 cells, one or more larger central cells surrounded by smaller cells, oil droplets present in all cells, 2-celled slime hairs with enlarged apical cell present in axils of leaves. Specialized asexual reproduction by caducous leaves or stems. Sexual condition dioicous, gametangia naked. Seta with foot, persistent, elongating prior to capsule maturation. Capsule erect, symmetric, well-defined neck region absent, stomates absent, dextrorsely spiralled at maturity, columella ephemeral, basifixed, not penetrating the archesporial tissue, peristome absent, dehiscence by longitudinal helical slit. Calyptra fugacious to rarely persistent, typically mitrate. Spores 3-radiate, slightly roughened to papillose. Genus 1, species 2 (2 in the flora): North America, se Asia (including Borneo). Takakiaceae are plants of cool, cold-temperate to arctic-alpine oceanic climates. They were first collected in the Himalayas by J. D. Hooker and placed in the hepatic genus Lepidozia as L. ceratophylla by W. Mitten. The consensus has been that it was a highly unusual liverwort with affinity to the Calobryales. Distinctive features include erect shoots arising from a stolon, terete leaves, sometimes fused at or near the base and thus of 2–4 segments, naked lateral gametangia, slime cells, oil droplets, and chromosome numbers of n = 4 or 5. -

Early Evolution of the Land Plant Circadian Clock
Research Early evolution of the land plant circadian clock Anna-Malin Linde1,2*, D. Magnus Eklund1,2*, Akane Kubota3*, Eric R. A. Pederson1,2, Karl Holm1,2, Niclas Gyllenstrand1,2, Ryuichi Nishihama3, Nils Cronberg4, Tomoaki Muranaka5, Tokitaka Oyama5, Takayuki Kohchi3 and Ulf Lagercrantz1,2 1Department of Plant Ecology and Evolution, Evolutionary Biology Centre, Uppsala University, Norbyv€agen 18D, SE-75236 Uppsala, Sweden; 2The Linnean Centre for Plant Biology in Uppsala, Uppsala, Sweden; 3Graduate School of Biostudies, Kyoto University, Kyoto 606-8502, Japan; 4Department of Biology, Lund University, Ecology Building, SE-22362 Lund, Sweden; 5Graduate School of Science, Kyoto University, Kyoto 606-8502, Japan Summary Author for correspondence: While angiosperm clocks can be described as an intricate network of interlocked transcrip- Ulf Lagercrantz tional feedback loops, clocks of green algae have been modelled as a loop of only two genes. Tel: +46(0)18 471 6418 To investigate the transition from a simple clock in algae to a complex one in angiosperms, we Email: [email protected] performed an inventory of circadian clock genes in bryophytes and charophytes. Additionally, Received: 14 December 2016 we performed functional characterization of putative core clock genes in the liverwort Accepted: 18 January 2017 Marchantia polymorpha and the hornwort Anthoceros agrestis. Phylogenetic construction was combined with studies of spatiotemporal expression patterns New Phytologist (2017) 216: 576–590 and analysis of M. polymorpha clock gene mutants. doi: 10.1111/nph.14487 Homologues to core clock genes identified in Arabidopsis were found not only in bryophytes but also in charophytes, albeit in fewer copies. Circadian rhythms were detected Key words: bryophyte, circadian clock, for most identified genes in M. -

NEW DATA ABOUT MOSSES on the SVALBARD GLACIERS Olga
NEW DATA ABOUT MOSSES ON THE SVALBARD GLACIERS Olga Belkina Polar-Alpine Botanical Garden-Institute, Kola Science Center of the Russian Academy of Sciences, Apatity, Murmansk Province, Russia; e-mail: [email protected] Rapid melting and retreat of glaciers in the Arctic is a cause of sustainable long‐ In Svalbard moss populations were found on 9 glaciers. In 2012, during re‐examination of the populations a few term existence of ablation zone on them. Sometimes these areas are the habitats In 2007 B.R.Mavlyudov collected one specimen on individuals of Bryum cryophilum Mårtensson and some plants of some mosses partly due to good availability of water and cryoconite Bertilbreen (Paludella squarrosa (Hedw.) Brid.) and of Sanionia uncinata were found among H. polare shoots in substratum. 14 species were found in this unusual habitat on Alaska and Iceland: some specimens on Austre Grønfjordbreen (Ceratodon some large cushions. Therefore the next stage of cushion Andreaea rupestris Hedw., Ceratodon purpureus (Hedw.) Brid., Ditrichum purpureus (Hedw.) Brid., Warnstorfia sarmentosa succession had begun –emergence of a di‐ and multi‐species flexicaule (Schwaegr.) Hampe, Pohlia nutans (Hedw.) Lindb., Polytrichum (Wahlenb.) Hedenäs, Sanionia uncinata (Hedw.) community. A similar process was observed earlier on the juniperinum Hedw. (Benninghoff, 1955), Racomitrium fasciculare (Hedw.) Brid. Loeske, Hygrohypnella polare (Lindb.) Ignatov & bone of a mammal that was lying on the same glacier. (=Codriophorus fascicularis (Hedw.) Bendarek‐Ochyra et Ochyra) (Shacklette, Ignatova.). Ceratodon purpureus settled in center of almost spherical 1966), Drepanocladus berggrenii (C.Jens.) Broth. (Heusser, 1972), Racomitrium In 2009 populations of two latter species were studied cushion of Sanionia uncinata on the both butt‐ends of the crispulum var. -

JUDD W.S. Et. Al. (2002) Plant Systematics: a Phylogenetic Approach. Chapter 7. an Overview of Green
UNCORRECTED PAGE PROOFS An Overview of Green Plant Phylogeny he word plant is commonly used to refer to any auto- trophic eukaryotic organism capable of converting light energy into chemical energy via the process of photosynthe- sis. More specifically, these organisms produce carbohydrates from carbon dioxide and water in the presence of chlorophyll inside of organelles called chloroplasts. Sometimes the term plant is extended to include autotrophic prokaryotic forms, especially the (eu)bacterial lineage known as the cyanobacteria (or blue- green algae). Many traditional botany textbooks even include the fungi, which differ dramatically in being heterotrophic eukaryotic organisms that enzymatically break down living or dead organic material and then absorb the simpler products. Fungi appear to be more closely related to animals, another lineage of heterotrophs characterized by eating other organisms and digesting them inter- nally. In this chapter we first briefly discuss the origin and evolution of several separately evolved plant lineages, both to acquaint you with these important branches of the tree of life and to help put the green plant lineage in broad phylogenetic perspective. We then focus attention on the evolution of green plants, emphasizing sev- eral critical transitions. Specifically, we concentrate on the origins of land plants (embryophytes), of vascular plants (tracheophytes), of 1 UNCORRECTED PAGE PROOFS 2 CHAPTER SEVEN seed plants (spermatophytes), and of flowering plants dons.” In some cases it is possible to abandon such (angiosperms). names entirely, but in others it is tempting to retain Although knowledge of fossil plants is critical to a them, either as common names for certain forms of orga- deep understanding of each of these shifts and some key nization (e.g., the “bryophytic” life cycle), or to refer to a fossils are mentioned, much of our discussion focuses on clade (e.g., applying “gymnosperms” to a hypothesized extant groups. -

Total of 10 Pages Only May Be Xeroxed
CENTRE FOR NEWFOUNDLAND STUDIES TOTAL OF 10 PAGES ONLY MAY BE XEROXED (Without Author's Permission) ,, l • ...J ..... The Disjunct Bryophyte Element of the Gulf of St. Lawrence Region: Glacial and Postglacial Dispersal and Migrational Histories By @Rene J. Belland B.Sc., M.Sc. A thesis submitted to the School of Graduate Studies in partial fulfilment of the requirements for the degree of Doctor of Philosophy Department of Biology Memorial University of Newfoundland December, 1Q84 St. John's Newfoundland Abstract The Gulf St. Lawrence region has a bryophyte flora of 698 species. Of these 267 (38%) are disjunct to this region from western North America, eastern Asia, or Europe. The Gulf of St. Lawrence and eastern North American distributions of the disjuncts were analysed and their possible migrational and dispersal histories during and after the Last Glaciation (Wisconsin) examined. Based on eastern North American distribution patterns, the disjuncts fell into 22 sub elements supporting five migrational/ dispersal histories or combinations of these : (1) migration from the south, (2) migration from the north, (3) migration from the west, (4) survival in refugia, and (5) introduction by man. The largest groups of disjuncts had eastern North American distributions supporting either survival of bryophytes in Wisconsin ice-free areas of the Gulf of St. Lawrence or postglacial migration to the Gulf from the south. About 26% of the disjuncts have complex histories and their distributions support two histories. These may have migrated to the Gulf from the west and/or north, or from the west and/or survived glaciation in Gulf ice-free areas. -

Extant Diversity of Bryophytes Emerged from Successive Post-Mesozoic Diversification Bursts
ARTICLE Received 20 Mar 2014 | Accepted 3 Sep 2014 | Published 27 Oct 2014 DOI: 10.1038/ncomms6134 Extant diversity of bryophytes emerged from successive post-Mesozoic diversification bursts B. Laenen1,2, B. Shaw3, H. Schneider4, B. Goffinet5, E. Paradis6,A.De´samore´1,2, J. Heinrichs7, J.C. Villarreal7, S.R. Gradstein8, S.F. McDaniel9, D.G. Long10, L.L. Forrest10, M.L. Hollingsworth10, B. Crandall-Stotler11, E.C. Davis9, J. Engel12, M. Von Konrat12, E.D. Cooper13, J. Patin˜o1, C.J. Cox14, A. Vanderpoorten1,* & A.J. Shaw3,* Unraveling the macroevolutionary history of bryophytes, which arose soon after the origin of land plants but exhibit substantially lower species richness than the more recently derived angiosperms, has been challenged by the scarce fossil record. Here we demonstrate that overall estimates of net species diversification are approximately half those reported in ferns and B30% those described for angiosperms. Nevertheless, statistical rate analyses on time- calibrated large-scale phylogenies reveal that mosses and liverworts underwent bursts of diversification since the mid-Mesozoic. The diversification rates further increase in specific lineages towards the Cenozoic to reach, in the most recently derived lineages, values that are comparable to those reported in angiosperms. This suggests that low diversification rates do not fully account for current patterns of bryophyte species richness, and we hypothesize that, as in gymnosperms, the low extant bryophyte species richness also results from massive extinctions. 1 Department of Conservation Biology and Evolution, Institute of Botany, University of Lie`ge, Lie`ge 4000, Belgium. 2 Institut fu¨r Systematische Botanik, University of Zu¨rich, Zu¨rich 8008, Switzerland. -
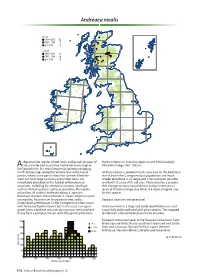
Andreaea Nivalis
Andreaea nivalis Britain 1990–2013 12 1950–1989 7 pre-1950 4 Ireland 1990–2013 0 1950–1989 0 pre-1950 0 characteristic species of wet rocks and gravels in areas of Nardia compressa, Scapania uliginosa and Pohlia ludwigii. Alate snow-lie and associated meltwater burns high in Altitudinal range: 880–1340 m. the Scottish hills. It is most frequently found on dripping, north-facing crags along the ‘cornice line’ at the top of Andreaea nivalis is abundant in its core area on the Ben Nevis corries, where snow persists into the summer. Here the massif and in the Cairngorms but populations are much moss can form large cushions and on Ben Nevis it is smaller elsewhere in its range and it has not been possible remarkably abundant in this habitat with numerous to refind it at some of its old sites. There must be a concern associates, including Deschampsia cespitosa, Saxifraga that changes in snow accumulation and persistence as a stellaris, Anthelia julacea, Lophozia opacifolia, Marsupella result of climate change may affect the more marginal sites sphacelata, M. stableri, Andreaea alpina, A. rupestris, for this species. Ditrichum zonatum, Kiaeria falcata, K. starkei, Polytrichastrum sexangulare, Racomitrium lanuginosum and, rarely, Dioicous; capsules are occasional. Oedipodium griffithianum. In the Cairngorms it often occurs with Andreaea frigida in burns but it also occurs on open Andreaea nivalis is a large and easily identifiable moss and gravel areas which are very wet during snow melt and here is not likely to be confused with other species. The mapped it may form a complex mosaic with Marsupella sphacelata, distribution is therefore believed to be accurate. -

THE ANDEAN SPECIES of PILEA by Ellsworth P. Killip INTRODUCTION
THE ANDEAN SPECIES OF PILEA By Ellsworth P. Killip INTRODUCTION Pilea is by far the largest genus of Urticaceae. Weddell in his final monograph 1 of the family recognized 159 species as valid, about 50 of which were said to occur in the Andes of South America. That these species had a very limited range of distribution was indicated by the fact that more than 30 were known from only a single collection and that only 9 of the others were in any sense widely distributed in the Andean countries. Two, P. microphyUa and P. pubescens, occurred throughout the American Tropics, and P. kyalina and P. dendrophUa extended eastward in South America. In the 50 years following the publication of this monograph only 10 species were described from the Andean region. Because of the large number of specimens of Pilea collected by expeditions to the Andes sponsored by the Smithsonian Institution, the New York Botanical Garden, the Gray Herbarium and the Arnold Arboretum of Harvard University, the Field Museum of Natural History, and the Academy of Natural Sciences, Philadelphia, which were submitted to me for identification but which could not be assigned to any known species, I began the preparation of a mono- graph of all the Andean species. This was completed in 1935 but unfortunately could not be printed in its entirety at that time. Instead, a greatly abridged paper was published,2 which contained a key to all the Andean species, descriptions of 30 new ones, and a few changes of name and of rank. In this there was no opportunity to present descriptions of the earlier species or to discuss their synonymy. -

Volume 1, Chapter 2-4: Bryophyta
Glime, J. M. 2017. Bryophyta - Takakiopsida. Chapt. 2-4. In: Glime, J. M. Bryophyte Ecology. Volume 1. Physiological Ecology. 2-4-1 Ebook sponsored by Michigan Technological University and the International Association of Bryologists. Last updated 9 April 2021 and available at <http://digitalcommons.mtu.edu/bryophyte-ecology/>. CHAPTER 2-4 BRYOPHYTA – TAKAKIOPSIDA TABLE OF CONTENTS Phylum Bryophyta .............................................................................................................................................. 2-4-2 Class Takakiopsida...................................................................................................................................... 2-4-2 Summary ........................................................................................................................................................... 2-4-10 Acknowledgments............................................................................................................................................. 2-4-10 Literature Cited ................................................................................................................................................. 2-4-10 2-4-2 Chapter 2-4: Bryophyta - Takakiopsida CHAPTER 2-4 BRYOPHYTA – TAKAKIOPSIDA Figure 1. Mt. Daisetsu from Kogan Spa, Hokkaido, Japan. The foggy peak of Mt. Daisetsu is the home of Takakia lepidozioides. Photo by Janice Glime. the Bryopsida, Andreaeopsida, and Sphagnopsida (Crum 1991). However, as more evidence from genetic and biochemical relationships -

Chapter 12 Productivity
Glime, J. M. 2017. Productivity. Chapt. 12. In: Glime, J. M. Bryophyte Ecology. Volume 1. Physiological Ecology. Ebook 12-1-1 sponsored by Michigan Technological University and the International Association of Bryologists. Last updated 18 July 2020 and available at <http://digitalcommons.mtu.edu/bryophyte-ecology/>. CHAPTER 12 PRODUCTIVITY TABLE OF CONTENTS Productivity .......................................................................................................................................................... 12-2 Ecological Factors ................................................................................................................................................ 12-2 Ability to Invade ........................................................................................................................................... 12-2 Niche Differences ......................................................................................................................................... 12-3 Growth ................................................................................................................................................................. 12-3 Growth Measurements .................................................................................................................................. 12-4 Annual Length Increase ................................................................................................................................ 12-8 Uncoupling ................................................................................................................................................... -

Morphology Supports the Setaphyte Hypothesis: Mosses Plus Liverworts Form a Natural Group
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/329947849 Morphology supports the setaphyte hypothesis: mosses plus liverworts form a natural group Article · December 2018 DOI: 10.11646/bde.40.2.1 CITATION READS 1 713 3 authors: Karen S Renzaglia Juan Carlos Villarreal Southern Illinois University Carbondale Laval University 139 PUBLICATIONS 3,181 CITATIONS 64 PUBLICATIONS 1,894 CITATIONS SEE PROFILE SEE PROFILE David J. Garbary St. Francis Xavier University 227 PUBLICATIONS 3,461 CITATIONS SEE PROFILE Some of the authors of this publication are also working on these related projects: Ultrastructure and pectin composition of guard cell walls View project Integrating red macroalgae into land-based marine finfish aquaculture View project All content following this page was uploaded by Karen S Renzaglia on 27 December 2018. The user has requested enhancement of the downloaded file. Bry. Div. Evo. 40 (2): 011–017 ISSN 2381-9677 (print edition) DIVERSITY & http://www.mapress.com/j/bde BRYOPHYTEEVOLUTION Copyright © 2018 Magnolia Press Article ISSN 2381-9685 (online edition) https://doi.org/10.11646/bde.40.2.1 Morphology supports the setaphyte hypothesis: mosses plus liverworts form a natural group KAREN S. RENZAGLIA1, JUAN CARLOS VILLARREAL A.2,3 & DAVID J. GARBARY4 1 Department of Plant Biology, Southern Illinois University, Carbondale, Illinois, USA 2Département de Biologie, Institut de Biologie Intégrative et des Systèmes (IBIS), Université Laval, Québec, Canada 3 Smithsonian Tropical Research Institute, Panama, Panama 4 St. Francis Xavier University, Antigonish, Nova Scotia, Canada The origin and early diversification of land plants is one of the major unresolved problems in evolutionary biology. -

BRYOPHYTES .Pdf
Diversity of Microbes and Cryptogams Bryophyta Geeta Asthana Department of Botany, University of Lucknow, Lucknow – 226007 India Date of submission: May 11, 2006 Version: English Significant Key words: Bryophyta, Hepaticopsida (Liverworts), Anthocerotopsida (Hornworts), , Bryopsida (Mosses). 1 Contents 1. Introduction • Definition & Systematic Position in the Plant Kingdom • Alternation of Generation • Life-cycle Pattern • Affinities with Algae and Pteridophytes • General Characters 2. Classification 3. Class – Hepaticopsida • General characters • Classification o Order – Calobryales o Order – Jungermanniales – Frullania o Order – Metzgeriales – Pellia o Order – Monocleales o Order – Sphaerocarpales o Order – Marchantiales – Marchantia 4. Class – Anthocerotopsida • General Characters • Classification o Order – Anthocerotales – Anthoceros 5. Class – Bryopsida • General Characters • Classification o Order – Sphagnales – Sphagnum o Order – Andreaeales – Andreaea o Order – Takakiales – Takakia o Order – Polytrichales – Pogonatum, Polytrichum o Order – Buxbaumiales – Buxbaumia o Order – Bryales – Funaria 6. References 2 Introduction Bryophytes are “Avascular Archegoniate Cryptogams” which constitute a large group of highly diversified plants. Systematic position in the plant kingdom The plant kingdom has been classified variously from time to time. The early systems of classification were mostly artificial in which the plants were grouped for the sake of convenience based on (observable) evident characters. Carolus Linnaeus (1753) classified