Novel SLCO2A1 Mutations Cause Gender-Differentiated Pachydermoperiostosis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Hypertrichosis Terminalis
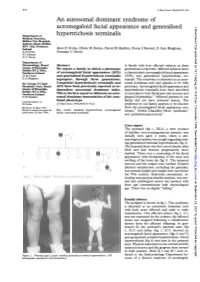
97297 Med Genet 1996;33:972-974 An autosomal dominant syndrome of acromegaloid facial appearance and generalised J Med Genet: first published as 10.1136/jmg.33.11.972 on 1 November 1996. Downloaded from Department of hypertrichosis terminalis Medical Genetics, Belfast City Hospital, Lisburn Road, Belfast BT9 7AB, Northern Ireland Alan D Irvine, Olivia M Dolan, David R Hadden, Fiona J Stewart, E Ann Bingham, A D Irvine Norman C Nevin F J Stewart N C Nevin Department of Dermatology, Royal Abstract A family with four affected subjects in three Group of Hospitals, Belfast BT12 6BA, We report a family in which a phenotype generations is reported. Affected subjects show Northern Ireland of acromegaloid facial appearance (AFA) a characteristic acromegaloid facial appearance 0 M Dolan and generalised hypertrichosis terminalis (AFA) and generalised hypertrichosis ter- E A Bingham segregates through three generations. minalis. The syndrome is inherited as an auto- Sir George E Clark Congenital hypertrichosis terminalis and somal dominant trait and appears to be fully Metabolic Unit, Royal AFA have been previously reported as in- penetrant. Acromegaloid facial appearance and Group of Hospitals, dependent autosomal dominant traits. hypertrichosis terminalis have been described Belfast BT12 6BA, Northern Ireland This is the first report to delineate an auto- in association with thickened oral mucosa and D R Hadden somal dominant transmission of the com- gingival hyperplasia.'"2 Affected patients in our Correspondence to: bined phenotype. family did not have intraoral lesions. The Dr Irvine. (JtMed Genet 1996;33:972-974) syndrome in our family appears to be discrete Received 14 May 1996 from the acromegaloid facial appearance syn- Revised version accepted for Key words: terminal hypertrichosis; acromegaloid drome,3 Gorlin-Chaudhry-Moss syndrome,4 publication 28 June 1996 facies; autosomal dominant. -

Digital Clubbing
Review Article Digital clubbing Malay Sarkar, D. M. Mahesh1, Irappa Madabhavi2 Department of Pulmonary Medicine, Indira Gandhi Medical College, Shimla, 1Department of Endocrinology, Jawaharlal Institute of Postgraduate Medical Education and Research, Pondicherry, 2Post Graduate Student (Medicine), Indira Gandhi Medical College, Shimla, India ABSTRACT Digital clubbing is an ancient and important clinical signs in medicine. Although clubbed fingers are mostly asymptomatic, it often predicts the presence of some dreaded underlying diseases. Its exact pathogenesis is not known, but platelet‑derived growth factor and vascular endothelial growth factor are recently incriminated in its causation. The association of digital clubbing with various disease processes and its clinical implications are discussed in this review. KEY WORDS: Cancer, digital clubbing, hypertrophic osteoarthropathy, megakaryocytes, vascular endothelial growth factor Address for correspondence: Dr. Malay Sarkar, Department of Pulmonary Medicine, Indira Gandhi Medical College, Shimla ‑ 171 001, India. E‑mail: [email protected] INTRODUCTION long bones and occasional painful joint enlargement. It was initially known as hypertrophic pulmonary Digital clubbing is characterized by a focal bulbous osteoarthropathy (HPOA) based on the fact that majority enlargement of the terminal segments of the fingers of cases of HOA are due to malignant thoracic tumors. and/or toes due to proliferation of connective tissue The term “pulmonary” was later abandoned as it was between nail matrix and the distal phalanx. It results in realized that the skeletal syndrome may occur in several increase in both anteroposterior and lateral diameter of non‑pulmonary diseases and even may occur without the nails.[1] Clubbed fingers are also known as watch‑glass any underlying illness. -

Pachydermoperiostosis in a Patient with Crohn's Disease: Treatment
IJMS Vol 43, No 1, January 2018 Case Report Pachydermoperiostosis in a Patient with Crohn’s Disease: Treatment and Literature Review Maryam Mobini1, MD; Abstract Ozra Akha1, MD; Hafez Fakheri2, MD; Pachydermoperiostosis (PDP) is a rare disorder characterized Hadi Majidi3, MD; by pachydermia, digital clubbing, periostitis, and an excess Sanam Fattahi4, MD of affected males. It is the primary form of hypertrophic osteoarthropathy (HOA) and there are some rare associations of PDP with other disorders. Here we describe a patient with 1Department of Internal Medicine, Diabetes Research Center, Faculty of Crohn’s disease associated with PDP. A 26-year-old man, who Medicine, Mazandaran University of was a known case of Crohn’s disease, referred with diffuse Medical Sciences, Sari, Iran; 2Department of Gastroenterology, swelling in the upper and lower limbs and cutis verticis gyrata Inflammatory Gut and Liver Research since 7 years ago. PDP was suspected and endocrinological Center, Mazandaran University of and radiological studies were conducted for the evaluation Medical Sciences, Sari, Iran; 3Department of Radiology, Orthopedic of underlying disease. He was prescribed celecoxib, low- Research Center, Faculty of Medicine, dose prednisolone, and pamidronate to control the swelling, Mazandaran University of Medical periostitis, azathiopurine, and mesalazine according to Sciences, Sari, Iran; 4Medical Student, Faculty of Medicine, gastrointestinal involvement. In conclusion, it is important to Mazandaran University of Medical identify this condition since a misdiagnosis might subject the Sciences, Sari, Iran patient to unnecessary investigations. Correspondence: Maryam Mobini, MD; Please cite this article as: Mobini M, Akha O, Fakheri H, Majidi H, Fattahi S. Imam Khomeini Hospital, Pachydermoperiostosis in a Patient with Crohn’s Disease: Treatment and Razi Street, Sari, Iran Literature Review. -

Pachydermoperiostosis (Touraine–Solente–Gole Syndrome): a Case
Joshi et al. Journal of Medical Case Reports (2019) 13:39 https://doi.org/10.1186/s13256-018-1961-z CASE REPORT Open Access Pachydermoperiostosis (Touraine–Solente– Gole syndrome): a case report Amir Joshi1* , Gaurav Nepal1, Yow Ka Shing2, Hari Prasad Panthi1 and Suman Baral1 Abstract Background: Pachydermoperiostosis (PDP) is a rare disorder characterized by clubbing of the fingers, thickening of the skin (pachyderma), and excessive sweating (hyperhidrosis). It typically appears during childhood or adolescence, often around the time of puberty, and progresses slowly. Clinical presentations of PDP can be confused with secondary hypertrophic osteoarthropathy, psoriatic arthritis, rheumatoid arthritis, thyroid acropachy, and acromegaly. Case presentation: A Mongolian male, aged 19 years, resident of a hilly district of Nepal, with history of consanguinity, presented to our outpatient department with chief complaints of pain and swelling in both hands and feet for 6 years. The pain was insidious in onset, throbbing in nature, and not relieved by over-the-counter medications. The patient also complained of profuse sweating, progressive enlargement of hands and feet, and gradual coarsening of facial features. On examination there were marked skin folds in the forehead, face, and eyelids. Clubbing and swelling of bilateral knee joints and ankle joints was also evident. He was subsequently investigated extensively for acromegaly. Insulin-like growth factor-1 level and oral glucose tolerance test were normal. Radiography of various bones showed -

Pachydermoperiostosis Mimicking Acromegaly: a Case Report
Turk J Rheumatol 2012;27(2):132-135 doi: 10.5606/tjr.2012.020 Case Report Pachydermoperiostosis Mimicking Acromegaly: A Case Report Akromegaliyi Taklit Eden Pakidermoperiostoz: Olgu Sunumu Mi-Hye KWON, Chung-Il JOUNG Division of Rheumatology, Konyang University Hospital, Daejeon, Korea Pachydermoperiostosis is a rare osteoarthroder-mopathic Pakidermoperiostoz, klinik ve radyografik tablosu disorder of which clinical and radiographic presentations akromegaliyi taklit edebilen bir osteoartrodermopatik may mimic those of acromegaly. In the evaluation of patients bozukluktur. Akromegaloid görünümlü hastalar with acromegaloid appearances, pachydermoperiostosis değerlendirilirken, pakidermoperiostoz ayırıcı tanı should be considered as a differential diagnosis. In this olarak akla getirilmelidir. Bu makalede el, ayak ve article, we present a 26-year-old man who was admitted ayak bileğinde altı aydır devam eden ağrılı şişlik ve with the complaints of painful swelling and stiffness of sertlik yakınması ile başvuran 26 yaşında erkek bir olgu hands, feet and ankles lasting for about six months. sunuldu. Key words: Arthralgia; pachydermoperiostosis; primary Anahtar sözcükler: Eklem ağrısı; pakidermoperiostoz; primer hypertrophic osteoarthropathy. hipertrofik osteoartropati. Pachydermoperiostosis (PDP), also referred to as most recent six month period, he had experienced primary hypertrophic osteoarthropathy (HOA), is a several episodes of lower leg edema which had rare hereditary disorder characterized by clubbing subsided completely within five days without any of the digits, thickening of the skin, and periosteal treatment, and joint pain in both wrists, elbows, changes in the long bones. When patients with PDP and ankles intermittently. Six years previously, he visit a hospital complaining of arthralgia, its clinical had undergone an operation for jaw misalignment and radiological presentation can be diagnostically and lantern jaw. -

Global Development Assistance for Adolescent Health from 2003 to 2015
Supplementary Online Content Li Z, Li M, Patton GC, Lu C. Global development assistance for adolescent health from 2003 to 2015. JAMA Netw Open. 2018;1(4):e181072. doi:10.1001/jamanetworkopen.2018.1072 eTable 1. List of Donor Countries Included in the CRS eTable 2. 132 Recipients in the CRS (According to the World Health Organization Regions) eTable 3. Key Words to Identify the Related Age Group (Adolescence) in the Creditor Reporting System eTable 4. CRS Purpose Name and Respective Fractions Allocated to Adolescent Health eTable 5. Definitions of DAAH on the Leading Causes of DALYs of Adolescent Health eTable 6. Key Words Used to Search for Projects on Skin and Subcutaneous Diseases in the Creditor Reporting System eTable 7. Key Words Used to Search for Road Injury Projects in the Creditor Reporting System eTable 8. Key Words Used to Search for HIV/AIDS Projects in the Creditor Reporting System eTable 9. Key Words Used to Search for Projects on Iron-Deficiency Anemia in the Creditor Reporting System eTable 10. Key Words Used to Search for Self-Harm Projects in the Creditor Reporting System eTable 11. Key Words Used to Search for Projects on Interpersonal Violence in the Creditor Reporting System eTable 12. Key Words Used to Search for Projects on Depressive Disorders in the Creditor Reporting System eTable 13. Key Words Used to Search for Projects on Lower Back and Neck Pain in the Creditor Reporting System eTable 14. Key Words Used to Search for Diarrheal Projects in the Creditor Reporting System eTable 15. Key Words Used to Search for Tuberculosis Projects in the Creditor Reporting System eTable 16. -

Acro-Osteolysis: a Complication of Jadassohn–Lewandowsky Syndrome
15 March 2005 Use of Articles in the Pachyonychia Congenita Bibliography The articles in the PC Bibliography may be restricted by copyright laws. These have been made available to you by PC Project for the exclusive use in teaching, scholar- ship or research regarding Pachyonychia Congenita. To the best of our understanding, in supplying this material to you we have followed the guidelines of Sec 107 regarding fair use of copyright materials. That section reads as follows: Sec. 107. - Limitations on exclusive rights: Fair use Notwithstanding the provisions of sections 106 and 106A, the fair use of a copyrighted work, including such use by reproduction in copies or phonorecords or by any other means specified by that section, for purposes such as criticism, comment, news reporting, teaching (including multiple copies for classroom use), scholarship, or research, is not an infringement of copyright. In determining whether the use made of a work in any particular case is a fair use the factors to be considered shall include - (1) the purpose and character of the use, including whether such use is of a commercial nature or is for nonprofit educational purposes; (2) the nature of the copyrighted work; (3) the amount and substantiality of the portion used in relation to the copyrighted work as a whole; and (4) the effect of the use upon the potential market for or value of the copyrighted work. The fact that a work is unpublished shall not itself bar a finding of fair use if such finding is made upon consideration of all the above factors. -

Cutis Verticis Gyrata in a Patient with Multiple Basal Cell
Journal of Mind and Medical Sciences Volume 3 | Issue 1 Article 10 2016 Cutis verticis gyrata in a patient with multiple basal cell carcinomas; case presentation and review of the literature Simona Roxana Georgescu Carol Davila University, Department of Dermatology and Venereology Maria Isabela Sârbu Carol Davila University, Department of Dermatology and Venereology, [email protected] Cristina Iulia Mitran Victor Babes Hospital, Department of Dermatology and Venereology Mădălina Irina Mitran Victor Babes Hospital, Department of Dermatology and Venereology Alice Rusu Victor Babes Hospital, Department of Dermatology and Venereology See next page for additional authors Follow this and additional works at: http://scholar.valpo.edu/jmms Part of the Dermatology Commons Recommended Citation Georgescu, Simona Roxana; Sârbu, Maria Isabela; Mitran, Cristina Iulia; Mitran, Mădălina Irina; Rusu, Alice; Benea, Vasile; and Tampa, Mircea (2016) "Cutis verticis gyrata in a patient with multiple basal cell carcinomas; case presentation and review of the literature," Journal of Mind and Medical Sciences: Vol. 3 : Iss. 1 , Article 10. Available at: http://scholar.valpo.edu/jmms/vol3/iss1/10 This Case Presentation is brought to you for free and open access by ValpoScholar. It has been accepted for inclusion in Journal of Mind and Medical Sciences by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. Cutis verticis gyrata in a patient with multiple basal cell carcinomas; case presentation and review of the literature Authors Simona Roxana Georgescu, Maria Isabela Sârbu, Cristina Iulia Mitran, Mădălina Irina Mitran, Alice Rusu, Vasile Benea, and Mircea Tampa This case presentation is available in Journal of Mind and Medical Sciences: http://scholar.valpo.edu/jmms/vol3/iss1/10 J Mind Med Sci. -

Pachydermoperiostosis: Scintigraphic, Thermographic, Plethysmographic, and Capillaroscopic Observations ADEL G
Ann Rheum Dis: first published as 10.1136/ard.42.1.98 on 1 February 1983. Downloaded from Annals ofthe Rheumatic Diseases, 1983, 42, 98-102 Case report Pachydermoperiostosis: scintigraphic, thermographic, plethysmographic, and capillaroscopic observations ADEL G. FAM, HYACINTH CHIN-SANG, AND COLIN A. RAMSAY From the Departments ofMedicine and Radiology, University of Toronto, and Rheumatic Disease Unit, Sunnybrook Medical Centre, Toronto, Ontario, Canada SUMMARY A unique case of pachydermoperiostosis, multiple epiphyseal dysplasia, and secondary osteoarthritis is described. An additional interesting feature was the presence of acroosteolysis of the distal phalanges of fingers and toes. Scintigraphic, thermographic, plethysmographic, and capillaroscopic studies suggested increased blood flow through clubbed fingers, raising the possibil- ity that this may play a role in the pathogenesis of pachydermoperiostosis. Pachydermoperiostosis (primary or idiopathic hyper- copyright. trophic osteoarthropathy) is a rare, often familial disease characterised by digital clubbing, cylindrical thickening of legs and forearms, sebaceous gland overactivity, hyperhidrosis, and symmetri- cal irregular periosteal ossification predominantly affecting the distal ends of long bones.`-3 Reported here is a unique case of pachydermo- periostosis and multiple epiphyseal dysplasia. Scinti- http://ard.bmj.com/ graphic, thermographic, plethysmographic, and capillaroscopic findings suggested increased blood flow through clubbed digits. An additional, interest- ;. ing feature was the presence of acroosteolysis of the distal phalanges of fingers and toes, a finding which has rarely been described in pachydermo- periostosis.` on September 29, 2021 by guest. Protected Case report A 56-year-old single Italian woman presented in 1979 with a history of clubbed fingers and toes associated with deformities offeet, knees, and elbows dating back to childhood. -

A Rare Cutis Verticis Gyrata Secondary to Cerebriform Intradermal Nevus: Case Report and Literature Review Weiliang Zeng1,2 and Lili Guo1,2*
Zeng and Guo BMC Surg (2021) 21:234 https://doi.org/10.1186/s12893-021-01229-9 CASE REPORT Open Access A rare cutis verticis gyrata secondary to cerebriform intradermal nevus: case report and literature review Weiliang Zeng1,2 and Lili Guo1,2* Abstract Background: Cutis verticis gyrate (CVG) is a rare morphologic syndrome that presents with hypertrophy and folding of the scalp. CVG can be classifed into three forms: primary essential, primary non-essential, and secondary. Cerebri- form intradermal nevus (CIN) is a rare cause of secondary CVG. We are here to report a rare case of CVG with an under- lying CIN and discuss the clinical course, treatment options, and critical screening guidelines for these patients. Case presentation: A 25 year-old male patient presented with a chief complaint of generalized hair loss in the scalp parietaloccipital region for a duration of 1 year and the hair loss area was occasionally accompanied by mild itching. The hair loss started gradually and worsened over time. In addition, he had scalp skin folds resembling the ridge and furrow of the cerebral cortex in the parietaloccipital region since birth. Physical examination revealed hypertrophy and formation of folds in the parietal-occipital area, forming 5 to 6 furrows and ridges. The size of the cerebriform mass was about 12.0 cm 8.5 cm, without other skin lesions. Difuse non-scarring hair loss was distributed on the posterior-parietal scalp, mid-parietal× scalp and superior-occipital scalp. The diseased tissue of the patient’s parietaloc- cipital area was excised under general anesthesia. -

Pachydermoperiostosis Vs. Acromegaly in a Patient with Cutis
erimenta xp l D E e r & m l a a t c o i l n o i Journal of Clinical & Experimental Glick et al., J Clin Exp Dermatol Res 2012, 3:1 l g y C f R DOI: 10.4172/2155-9554.1000142 o e l ISSN: 2155-9554 s a e n a r r u c o h J Dermatology Research Case Report Open Access Pachydermoperiostosis vs. Acromegaly in a Patient with Cutis Verticis Gyrata Jaimie Glick, Ravneet Ruby Kaur* and Gina Taylor Department of Dermatology, State University of New York Downstate Medical Center, USA Abstract Cutis Verticis Gyrata (CVG) is a rare condition associated with thickening of the skin folds of the scalp and forehead that resembles the appearance of gyri of the brain cortex. These skin lesions can lead to local skin infections as well as cosmetic complaints. Primary CVG has no obvious cause and is often associated with neurological conditions while secondary CVG is due to inflammation or infiltration of the scalp and is associated with endocrine or neoplastic conditions including acromegaly and pachydermoperiostosis. We discuss a complicated case of a 34 year-old male with cutis verticis gyrata presenting simultaneously with hypertrophic osteoarthropathy and a pituitary macroadenoma. The patient was initially suspected of having acromegaly secondary to the pituitary adenoma but further workup showed normal insulin growth factor-1 and growth hormone levels inconsistent with acromegaly. Subsequent workup was consistent with a diagnosis of the complete form of primary hypertrophic osteoarthropathy also known as pachydermoperiostosis. Introduction He had noted swelling with enlargement of his hands and feet over the last few years. -

Jennifer a Cafardi the Manual of Dermatology 2012
The Manual of Dermatology Jennifer A. Cafardi The Manual of Dermatology Jennifer A. Cafardi, MD, FAAD Assistant Professor of Dermatology University of Alabama at Birmingham Birmingham, Alabama, USA [email protected] ISBN 978-1-4614-0937-3 e-ISBN 978-1-4614-0938-0 DOI 10.1007/978-1-4614-0938-0 Springer New York Dordrecht Heidelberg London Library of Congress Control Number: 2011940426 © Springer Science+Business Media, LLC 2012 All rights reserved. This work may not be translated or copied in whole or in part without the written permission of the publisher (Springer Science+Business Media, LLC, 233 Spring Street, New York, NY 10013, USA), except for brief excerpts in connection with reviews or scholarly analysis. Use in connection with any form of information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology now known or hereafter developed is forbidden. The use in this publication of trade names, trademarks, service marks, and similar terms, even if they are not identifi ed as such, is not to be taken as an expression of opinion as to whether or not they are subject to proprietary rights. While the advice and information in this book are believed to be true and accurate at the date of going to press, neither the authors nor the editors nor the publisher can accept any legal responsibility for any errors or omissions that may be made. The publisher makes no warranty, express or implied, with respect to the material contained herein. Printed on acid-free paper Springer is part of Springer Science+Business Media (www.springer.com) Notice Dermatology is an evolving fi eld of medicine.