The First Dinosaur Egg Was Soft
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
![[PDF] Dinosaur Eggshell from the Red Sandstone Group of Tanzania](https://docslib.b-cdn.net/cover/9168/pdf-dinosaur-eggshell-from-the-red-sandstone-group-of-tanzania-179168.webp)
[PDF] Dinosaur Eggshell from the Red Sandstone Group of Tanzania
Journal of Vertebrate Paleontology 24(2):494±497, June 2004 q 2004 by the Society of Vertebrate Paleontology NOTE DINOSAUR EGGSHELL FROM THE RED SANDSTONE GROUP OF TANZANIA MICHAEL D. GOTTFRIED1, PATRICK M. O'CONNOR2, FRANKIE D. JACKSON3, ERIC M. ROBERTS4, and REMEGIUS CHAMI5, 1Mich- igan State University Museum, East Lansing, Michigan, 48824, [email protected]; 2College of Osteopathic Medicine, Ohio University, Athens, Ohio, 45701; 3Department of Earth Sciences, Montana State University, Bozeman, Montana, 59717; 4Department of Geology and Geophysics, University of Utah, Salt Lake City, Utah, 84112, 5Antiquities Unit, P.O. Box 2280, Dar es Salaam, Tanzania Investigations over the last several decades at Gondwanan Mesozoic Although the age of the Red Sandstone Group is poorly understood (see localities have signi®cantly expanded our knowledge of the diversity Damblon et al., 1998), a Cretaceous age is suggested at this site based and distribution of Southern Hemisphere dinosaurs. These records are on (1) the overall composition of the fauna, which includes titanosaurid? primarily based on skeletal remains, but included among them are in- sauropods and both avian and nonavian theropods, as well as osteo- stances of preserved eggshell, notably from Argentina (e.g., Calvo et glossomorph ®shes, and (2) the possibility that these deposits may be al., 1997; Chiappe et al., 1998) and India (e.g., Khosla and Sahni, 1995). approximately coeval with the Cretaceous dinosaur beds of Malawi (Ja- In general, however, dinosaur eggshell is relatively poorly known from cobs et al., 1990), which lie ca. 200 km southeast of the Mbeya region. Gondwana, and from Africa in particular. -

Implications for the Evolution of the North Alpine Foreland Basin During the Miocene Climate Optimum
Vertebrate microfossils from the Upper Freshwater Molasse in the Swiss Molasse Basin: Implications for the evolution of the North Alpine Foreland Basin during the Miocene Climate Optimum Authors: Jürg Jost, Daniel Kälin, Saskia Börner, Davit Vasilyan, Daniel Lawver, & Bettina Reichenbacher NOTICE: this is the author’s version of a work that was accepted for publication in Palaeogeography, Palaeoclimatology, Palaeoecology. Changes resulting from the publishing process, such as peer review, editing, corrections, structural formatting, and other quality control mechanisms may not be reflected in this document. Changes may have been made to this work since it was submitted for publication. A definitive version was subsequently published in Palaeogeography, Palaeoclimatology, Palaeoecology, [Vol# 426, (May 15, 2015)] DOI# 10.1016/j.palaeo.2015.02.028 Jost, Jurg, Daniel Kalin, Saskia Borner, Davit Vasilyan, Daniel Lawver, and Bettina Reichenbacher. "Vertebrate microfossils from the Upper Freshwater Molasse in the Swiss Molasse Basin: Implications for the evolution of the North Alpine Foreland Basin during the Miocene Climate Optimum." Palaeogeography, Palaeoclimatology, Palaeoecology 426 (May 2015): 22-33. DOI: https://dx.doi.org/10.1016/j.palaeo.2015.02.028. Made available through Montana State University’s ScholarWorks scholarworks.montana.edu Vertebrate microfossils from the Upper Freshwater Molasse in the Swiss Molasse Basin: Implications for the evolution of the North Alpine Foreland Basin during the Miocene Climate Optimum a b c d e c Jürg Jost , Daniel Kälin , Saskia Börner , Davit Vasilyan , Daniel Lawver , Bettina Reichenbacher a Bärenhubelstraße 10, CH-4800 Zofingen, Switzerland b Bundesamt für Landestopographie swisstopo, Geologische Landesaufnahme, Seftigenstrasse 264, 3084 Wabern, Switzerland c Department of Earth and Environmental Sciences, Section on Palaeontology and Geobiology, Ludwig-Maximilians-University, Richard-Wagner Str. -

Abstract Book.Pdf
Welcome! Welcome to the VI Symposium on Dinosaur Eggs and Babies, the return of this periodic gathering to the Iberian Peninsula, when it hatched eighteen years ago. From the slopes of the Pyrenees, we have followed the first steps of dinosaurs through France, Argentina, the United States and China. Today, we come back and see the coast where the first theropod embryos were discovered twenty years ago. Since the end of the last century, Paleoology, much like other branches of palaeontology, has evolved thanks to the advance of new methodologies and analytical tools, becoming a progressively more interdisciplinary area of knowledge. Dinosaur babies and embryos, rare findings back when these meetings started, seem to be everywhere now that we learn to look for them under the light of the microscope. New astonishing specimens allow us to understand how Mesozoic dinosaurs mate and reproduce. Oology, our parent discipline in the modern world, has made great advances in understanding the form and function of the egg, and its applications on poultry industry are countless. More than thirty contributions evidence that our field remains small but alive and healthy. We hope that you find in this Symposium an opportunity to share knowledge and open new lines of collaboration. And do not forget to enjoy your stay in Portugal. The host committee CONTENTS How to get to the FCT 6 Acknowledgements 10 PROGRAM 11 ABSTRACTS 14 THE FIRST ORNITHOMIMID EMBRYO IN A SHELL WITH A SINGLE STRUCTURAL LAYER: A CHALLENGE TO ORTHODOXY 15 Araújo R., Lamb J., Atkinson P., Martins R. M. S., Polcyn M.J., Fernandez V. -

Strange Tadpoles from the Lower Miocene of Turkey: Is Paedogenesis Possible in Anurans?
Strange tadpoles from the lower Miocene of Turkey: Is paedogenesis possible in anurans? ALAIN DUBOIS, STÉPHANE GROSJEAN, and JEAN−CLAUDE PAICHELER Dubois, A., Grosjean, S., and Paicheler, J.−C. 2010. Strange tadpoles from the lower Miocene of Turkey: Is paedogenesis possible in anurans? Acta Palaeontologica Polonica 55 (1): 43–55. Fossil material from the lower Miocene collected in the basin lake of Beşkonak (Turkey) included 19 slabs showing 19 amphibian anuran tadpoles of rather large size, at Gosner stages 36–38. These well preserved specimens show many mor− phological and skeletal characters. They are here tentatively referred to the genus Pelobates. Two of these tadpoles show an unusual group of black roundish spots in the abdominal region, and a third similar group of spots is present in another slab but we were unable to state if it was associated with a tadpole or not. Several hypotheses can be proposed to account for these structures: artefacts; intestinal content (seeds; inert, bacterial or fungal aggregations; eggs); internal or external parasites; diseases; eggs produced by the tadpole. The latter hypothesis is discussed in detail and is shown to be unlikely for several reasons. However, in the improbable case where these spots would correspond to eggs, this would be the first reported case of natural paedogenesis in anurans, a phenomenon which has been so far considered impossible mostly for anatomical reasons (e.g., absence of space in the abdominal cavity). Key words: Amphibia, Anura, egg, fossil, paedogenesis, tadpole, Miocene, Turkey. Alain Dubois [[email protected]] and Stéphane Grosjean [[email protected]], Département de Sytématique & Evolu− tion, Muséum National d’Histoire Naturelle, UMR 7205 Origine, Structure et Evolution de la Biodiversité, Reptiles et Amphibiens, Case 30, 25 rue Cuvier, 75005 Paris, France; Jean−Claude Paicheler [[email protected]], Laboratoire de Paléoparasitologie, UFR de Pharmacie, 51 rue Cognacq Jay, 51096 Reims Cedex, France. -

A Stable Isotopic Investigation of Resource Partitioning Among Neosauropod Dinosaurs of the Upper Jurassic Morrison Formation
A stable isotopic investigation of resource partitioning among neosauropod dinosaurs of the Upper Jurassic Morrison Formation Benjamin T. Breeden, III SID: 110305422 [email protected] GEOL394H University of Maryland, College Park, Department of Geology 29 April 2011 Advisors: Dr. Thomas R. Holtz1, Jr., Dr. Alan Jay Kaufman1, and Dr. Matthew T. Carrano2 1: University of Maryland, College Park, Department of Geology 2: National Museum of Natural History, Department of Paleobiology ABSTRACT For more than a century, morphological studies have been used to attempt to understand the partitioning of resources in the Morrison Fauna, particularly between members of the two major clades of neosauropod (long-necked, megaherbivorous) dinosaurs: Diplodocidae and Macronaria. While it is generally accepted that most macronarians fed 3-5m above the ground, the feeding habits of diplodocids are somewhat more enigmatic; it is not clear whether diplodocids fed higher or lower than macronarians. While many studies exploring sauropod resource portioning have focused on differences in the morphologies of the two groups, few have utilized geochemical evidence. Stable isotope geochemistry has become an increasingly common and reliable means of investigating paleoecological questions, and due to the resistance of tooth enamel to diagenetic alteration, fossil teeth can provide invaluable paleoecological and behavioral data that would be otherwise unobtainable. Studies in the Ituri Rainforest in the Democratic Republic of the Congo, have shown that stable isotope ratios measured in the teeth of herbivores reflect the heights at which these animals fed in the forest due to isotopic variation in plants with height caused by differences in humidity at the forest floor and the top of the forest exposed to the atmosphere. -

Cairanoolithus: a Large Egg for a Small Dinosaur with Wide Hips
20/11/2020 <em>Cairanoolithus</em>: a Large Egg for a Small Dinosaur With Wide Hips - UABDivulga Barcelona Research & Innovation 02/2015 Cairanoolithus: a Large Egg for a Small Dinosaur With Wide Hips The study of the microstructure of the eggshell of Cairanoolithus conducted by Albert G. Sellés and Angel Galobart, both researchers at the Catalan Institute of Palaeontology Miquel Crusafont (ICP), reveals that this egg type does not belong to sauropod dinosaurs but to ankylosaurs, and probably to Struthiosaurus, a genus of armored dinosaurs. The finding would represent the first description of thyreophora eggs (a group that includes ankylosauria and stegosauria) in the world. It is an arduous task for paleontologists to assign the fossil eggs they found in excavations to a certain dinosaur species or group of dinosaurs, especially because the lack of bones of the parents in the nests and the low probability to find embryonic remains that would help in the taxonomic assignation of the eggs. Thus, we often do not known who laid a certain fossil egg. These limitations have led scientist to design a specific system for naming and classifying fossil eggs, using concepts such as "oogenera" and "ooespecies" for its phylogenetic classification. Cairanoolithus is an oogenus discovered and described in the early 90’s. It takes its name from the first place where it was found, a site near La Cairanne, a small town from southeastern France. From then on, Cairanoolithus has been found in 25 different sites. These eggs are between 72.2 and 71.4 million years old, they are large (over 15 cm in diameter) and have a rounded shape. -

From the Adamantina Formation, Bauru Group, Upper Cretaceous of Brazil and the Phylogenetic Relationships of Aeolosaurini
Zootaxa 3085: 1–33 (2011) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2011 · Magnolia Press ISSN 1175-5334 (online edition) A new sauropod (Macronaria, Titanosauria) from the Adamantina Formation, Bauru Group, Upper Cretaceous of Brazil and the phylogenetic relationships of Aeolosaurini RODRIGO M. SANTUCCI1 & ANTONIO C. DE ARRUDA-CAMPOS2 1Universidade de Brasília - Faculdade UnB Planaltina, Brasília-DF, 73300-000, Brazil. E-mail: [email protected] 2Museu de Paleontologia de Monte Alto, Praça do Centenário, s/n. Monte Alto-SP, 15910-000, Brazil. E-mail: [email protected] Table of contents Abstract . 1 Introduction . 2 Historical background . 2 Geological setting . 4 Systematic Palaeontology . 4 DINOSAURIA Owen, 1842 . 4 SAURISCHIA Seeley, 1887 . 4 SAUROPODA Marsh, 1878 . 4 MACRONARIA Wilson and Sereno, 1998. 4 TITANOSAURIFORMES Salgado, Coria and Calvo, 1997b. 4 TITANOSAURIA Bonaparte and Coria, 1993 . 4 AEOLOSAURINI Franco-Rosas, Salgado, Rosas and Carvalho, 2004 . 5 Aeolosaurus Powell, 1987 . 5 Aeolosaurus rionegrinus Powell, 1987 . 5 Aeolosaurus maximus sp. nov. 6 Phylogenetic analysis . 17 Comparison and discussion . 19 Conclusions . 25 Acknowledgements . 25 References . 26 APPENDIX 1. 29 APPENDIX 2. 29 Abstract Remains of a new titanosaur, Aeolosaurus maximus sp. nov., from the Adamantina Formation (Upper Cretaceous), Bauru Group, São Paulo State of Brazil are described. The new species is represented by a single partially articulated skeleton and is characterized by having a well-developed posterior protuberance below the articular area on the anterior and middle haemal arches and a lateral bulge on the distal portion of the articular process of the mid-posterior haemal arches. It shares with other Aeolosaurus species the presence of prezygapophyses curved downward on anterior caudal vertebrae and hae- mal arches with double articular facets set in a concave posterodorsal surface. -

High European Sauropod Dinosaur Diversity During Jurassic–Cretaceous Transition in Riodeva (Teruel, Spain)
CORE Metadata, citation and similar papers at core.ac.uk Provided by RERO DOC Digital Library [Palaeontology, Vol. 52, Part 5, 2009, pp. 1009–1027] HIGH EUROPEAN SAUROPOD DINOSAUR DIVERSITY DURING JURASSIC–CRETACEOUS TRANSITION IN RIODEVA (TERUEL, SPAIN) by RAFAEL ROYO-TORRES*, ALBERTO COBOS*, LUIS LUQUE*, AINARA ABERASTURI*, , EDUARDO ESPI´LEZ*, IGNACIO FIERRO*, ANA GONZA´ LEZ*, LUIS MAMPEL* and LUIS ALCALA´ * *Fundacio´n Conjunto Paleontolo´gico de Teruel-Dino´polis. Avda. Sagunto s ⁄ n. E-44002 Teruel, Spain; e-mail: [email protected] Escuela Taller de Restauracio´n Paleontolo´gica II del Gobierno de Arago´n. Avda. Sagunto s ⁄ n. E-44002 Teruel, Spain Typescript received 13 December 2007; accepted in revised form 3 November 2008 Abstract: Up to now, more than 40 dinosaur sites have (CPT-1074) referring to the Diplodocidae clade. New been found in the latest Jurassic – earliest Cretaceous remains from the RD-28, RD-41 and RD-43 sites, of the sedimentary outcrops (Villar del Arzobispo Formation) of same age, among which there are caudal vertebrae, are Riodeva (Iberian Range, Spain). Those already excavated, assigned to Macronaria. New sauropod footprints from the as well as other findings, provide a large and diverse Villar del Arzobispo Formation complete the extraordinary number of sauropod remains, suggesting a great diversity sauropod record coming to light in the area. The inclusion for this group in the Iberian Peninsula during this time. of other sauropods from different contemporaneous expo- Vertebrae and ischial remains from Riodevan site RD-13 sures in Teruel within the Turiasauria clade adds new evi- are assigned to Turiasaurus riodevensis (a species described dence of a great diversity of sauropods in Iberia during in RD-10, Barrihonda site), which is part of the the Jurassic–Cretaceous transition. -

Paleontological Discoveries in the Chorrillo Formation (Upper Campanian-Lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina
Rev. Mus. Argentino Cienc. Nat., n.s. 21(2): 217-293, 2019 ISSN 1514-5158 (impresa) ISSN 1853-0400 (en línea) Paleontological discoveries in the Chorrillo Formation (upper Campanian-lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina Fernando. E. NOVAS1,2, Federico. L. AGNOLIN1,2,3, Sebastián ROZADILLA1,2, Alexis M. ARANCIAGA-ROLANDO1,2, Federico BRISSON-EGLI1,2, Matias J. MOTTA1,2, Mauricio CERRONI1,2, Martín D. EZCURRA2,5, Agustín G. MARTINELLI2,5, Julia S. D´ANGELO1,2, Gerardo ALVAREZ-HERRERA1, Adriel R. GENTIL1,2, Sergio BOGAN3, Nicolás R. CHIMENTO1,2, Jordi A. GARCÍA-MARSÀ1,2, Gastón LO COCO1,2, Sergio E. MIQUEL2,4, Fátima F. BRITO4, Ezequiel I. VERA2,6, 7, Valeria S. PEREZ LOINAZE2,6 , Mariela S. FERNÁNDEZ8 & Leonardo SALGADO2,9 1 Laboratorio de Anatomía Comparada y Evolución de los Vertebrados. Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina - fernovas@yahoo. com.ar. 2 Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina. 3 Fundación de Historia Natural “Felix de Azara”, Universidad Maimonides, Hidalgo 775, C1405BDB Buenos Aires, Argentina. 4 Laboratorio de Malacología terrestre. División Invertebrados Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina. 5 Sección Paleontología de Vertebrados. Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina. 6 División Paleobotánica. Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina. 7 Área de Paleontología. Departamento de Geología, Universidad de Buenos Aires, Pabellón 2, Ciudad Universitaria (C1428EGA) Buenos Aires, Argentina. 8 Instituto de Investigaciones en Biodiversidad y Medioambiente (CONICET-INIBIOMA), Quintral 1250, 8400 San Carlos de Bariloche, Río Negro, Argentina. -
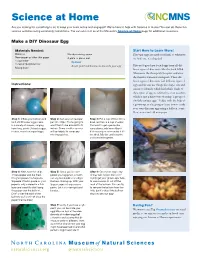
Make a DIY Dinosaur Egg (PDF)
Science at Home Are you looking for something to do to keep your brain active and engaged? We’re here to help with Science at Home! You can do these fun science activities using commonly found items. You can also visit us at the Museum’s Science at Home page for additional resources. Make a DIY Dinosaur Egg Materials Needed: Start Here to Learn More! Balloons Wooden mixing spoon Dinosaur eggs are rarely fossilized, so whenever Newspaper or other thin paper A plate or place-mat we find one, it’s a big deal. 1 cup water Optional 1 cup all-purpose flour Acrylic paint and brushes to decorate your egg Paleontologists have found eggs from all dif- Mixing bowl ferent types of dinosaurs, like the duck-billed Maiasaura, the theropod Oviraptor, and even the massive titanosaur sauropods. These dif- ferent types of dinosaurs laid different types of Instructions: eggs and we can use things like shape, size and texture to identify which laid which. Each of these types of egg are referred to as an ootaxon, which is just a fancy way of saying “a group of similarly unique eggs.” Today, with the help of a grown-up, you’re going to learn how to make your own dinosaur eggs using a balloon, some flour, water and old newspaper. Step 1: Inflate your balloon and Step 2: Cut up your newspa- Step 3: Put a cup of flour into a tie it off. Dinosaur eggs came per into strips. You’re going to bowl, and mix in a cup of water. -

Map 4-7-4 City Planning of Tongling Mun/Ipality , I4-2-5
E685 Vol. 4 World Bank Financed Project Anhui Expressway Project II Public Disclosure Authorized Public Disclosure Authorized Tongling-Tangkou Expressway Project Environment Assessment Report (Third Edition) Public Disclosure Authorized World Bank Financed Project Office of Anhui Provincial Communications Dzpartment Public Disclosure Authorized Dec. 2002 FILE COPY Tongling-Tangkou highway project EIA CONTENTS Chapter I Introduction ............................................ I 1.1 Project Background ............................................ I 1.2 Progress of EA ........................................... .1 1.3 Purpose of EA ............................................. 2 1.4 Bases of Assessment ........................................... .2 1.5 Technical Process for EA ....... ..................................... 3 1.6 Scope of Assessment ............................................ 5 1.7 Methodology ............................................ 5 1.8 Applicable Standards ............................................ 5 1.8.1 Ambient Air Quality Standards ........................................... 6 1.8.2 Environmental Noise Standards .................. ......................... 6 1.8.3 Surface Water Quality Standards ............................................ 6 Chapter 2 Environmental Assessment Team ......... .................................. 8 Chapter 3 Project Description ............................................ 10 3.1 Direct Project Benefit Area ........................................... 10 3.2 Geographical Location ..... -

ISPH-PROGRAM-And-ABSTRACT-BOOK.Pdf
- ISPH 2015 logo (front cover) designed by Jasmina Wiemann. The logo highlights several aspects well suited for the Bonn, 2015 meeting. The dwarf sauropod dinosaur, Europasaurus, stands in front of a histology-filled silhouette of the main dome of Poppelsdorf Palace, the main venue for ISPH 2015. This Late Jurassic sauropod is a fitting representative, as it was discovered in Lower Saxony, Germany, and its dwarf status was verified with histological investigations. - Cover, program and abstract book designed by Aurore Canoville and Jessica Mitchell. - Program and abstract book editors: Aurore Canoville, Jessica Mitchell, Koen Stein, Dorota Konietzko-Meier, Elzbieta Teschner, Anneke van Heteren, and P. Martin Sander. ISPH 2015 – Bonn, Germany ISPH 2015 – Bonn, Germany Table of Contents TABLE OF CONTENTS Symposium Organizers and Acknowledgements ...................................................... 4 Welcome Address ........................................................................................................ 5 Program ........................................................................................................................ 6 Main Events ........................................................................................................ 6 Venues ................................................................................................................ 8 Scientific Sessions / Oral Presentations ........................................................... 11 Scientific Sessions / List of Posters .................................................................