[PDF] Dinosaur Eggshell from the Red Sandstone Group of Tanzania
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Morrison Formation 37 Cretaceous System 48 Cloverly Formation 48 Sykes Mountain Formation 51 Thermopolis Shale 55 Mowry Shale 56
THE STRUCTURAL AND STRATIGRAPHIC FRAMEWORK OF THE WARM SPRINGS RANCH AREA, HOT SPRINGS COUNTY, WYOMING By CHRISTOPHER JAY CARSON Bachelor of Science Oklahoma State University 1998 Submitted to the Faculty of the Graduate College of the Oklahoma State University in partial fulfillment of the requirements for the Degree of MASTER OF SCIENCE July, 2000 THE STRUCTURAL AND STRATIGRAPHIC FRAMEWORK OF THE WARM SPRINGS RANCH AREA, HOT SPRINGS COUNTY, WYOMING Thesis Approved: Thesis Advisor ~~L. ... ~. ----'-"'-....D~e~e:.-g-e----- II ACKNOWLEDGEMENTS I wish to express appreciation to my advisor Dr. Arthur Cleaves for providing me with the opportunity to compile this thesis, and his help carrying out the fieldwork portion of the thesis. My sincere appreciation is extended to my advisory committee members: Dr. Stan Paxton, Dr. Gary Stewart, and Mr. David Schmude. I wish to thank Mr. Schmude especially for the great deal of personal effort he put forth toward the completion of this thesis. His efforts included financial, and time contributions, along with invaluable injections of enthusiasm, advice, and friendship. I extend my most sincere thank you to Dr. Burkhard Pohl, The Big Hom Basin Foundation, and the Wyoming Dinosaur Center. Without whose input and financial support this thesis would not have been possible. In conjunction I would like to thank the staff of the Wyoming Dinosaur Center for the great deal of help that I received during my stay in Thermopolis. Finally I wish to thank my friends and family. To my friends who have pursued this process before me, and with me; thank you very much. -

Communicating Science for Conservation
Communicating Science for Conservation Vol. XXXIV, No. 5, May 2019 ISSN 0971-6378 (Print); 0973-2543 (Online) Magazine of Zoo Outreach Organization www.zoosprint.zooreach.org Communicating science for conservation Vol. XXXIV, No. 5, May 2019 ISSN 0971-6378 (Print); 0973-2543 (Online) Contents Fantastic Facts Amazing Spider Facts, Part-I, -- Latha G Ravikumar, Pp. 1–4 Jottings These plants are thriving in extreme cliff conditions – here’s how -- Vidya Mary George, Pp. 5–6 Instagram Instagram images, P. 7 Highlight Review of dinosaur egg fossils from Gujarat State, India -- Raju V. Vyas, Pp. 8–17 Bird-o-soar Protective tactics of the Red-wattled Lapwing -- B. Ravichandran, Pp. 18–19 Reptile Rap Distribution record of Python bivittatus in Amangarh Tiger Reserve, Uttar Pradesh, India -- Prajakta Hushangabadkar, Meraj Anwar & Shariq Shafi, Pp. 20–22 Bugs R All Red Helen of the evergreen forests found for the first time in western West Bengal -- Kalyan Mukherjee, Pp. 23–24 Record of the aquatic leech Poecelobdella manilensis from Porur Lake, Chennai -- Muthukaruppan Gobi & Richard Thilagaraj, Pp. 25–26 Mammal Tales Colour abnormality of Macaca radiata at Grizzled Giant Squirrel Wildlife Sanctuary, Tamil Nadu, India -- R. Deepan, B. Vickram & A. Samson, Pp. 27–28 Field Report Tata Steel Zoological Park, Jamshedpur, Jharkhand - Residential Nature Camp on Awareness on Biodiversity & Willdife Conservation and Earth Day 2019 celebrations -- Seem Rani, Pp. 29–31 Workshop on Climate Change and Wildlife at Institute of Forestry, Pokhara, Nepal -- Prabhat Kiran Bhattarai, Pp. 32–34 Workshop on House Sparrow: Concern and Conservation in District Institute of Education and Training (DIET)-Lalitpur, Uttar Pradesh -- Akhilesh Kumar, Sonika Kushwaha & Pushpendra Singh Chauhan, Pp. -

Cáscaras De Huevos De Dinosaurios De La Formación Allen (Campaniano-Maastrichtiano), En Salitral Moreno, Provincia De Río Negro, Argentina
AMEGHINIANA (Rev. Asoc. Paleontol. Argent.) - 43 (3): 513-528. Buenos Aires, 30-9-2006 ISSN 0002-7014 Cáscaras de huevos de dinosaurios de la Formación Allen (Campaniano-Maastrichtiano), en Salitral Moreno, provincia de Río Negro, Argentina María Edith SIMÓN1 Abstract. DINOSAUR EGGSHELLS FROM THE ALLEN FORMATION (CAMPANIAN-MAASTRICHTIAN), AT SALITRAL MORENO, RÍO NEGRO PROVINCE, ARGENTINA. A detailed description of two morphotypes of dinosaur eggshells from Salitral Moreno, Rio Negro Province (Allen Formation, Upper Cretaceous) is presented. One of them is described as Patagoolithus salitralensis, a new oogenus et oospecies nov. belonging to the Megaloolithidae, and the second is referred as ?Faveoloolithidae, oogenus et oospecies indet. A detailed ultrastructural description of both morphotypes is provided, as well as, the chemical and mineralogical analysis of ?Faveoloolithidae oogen. et oospec. indet. In the studied sections, three eggshell-bearing levels were recognized. In the lower one (NH1) Patagoolithus salitralensis is associated to a diverse fauna of di- nosaurs, chelonians and invertebrates, and the few shell fragments are dispersed, suggesting deposition in an alluvial plain or disintegrated eggs. In NH2 and NH3, only eggshells of ?Faveoloolithidae oogenus et oospecies indet. were found. NH2 shows lateral variations in the inferred degree of transport and den- sity of the shell fragments in different sections. NH3 bears small shell fragments of the same morphotype decreasing in number towards the top. NH1 and NH3 bear rizolithes and alveolar structures, characteris- tic of calcretes, which could be indicative of palaeosoils. Resumen. En el presente trabajo se describen en detalle dos morfotipos de cáscaras de huevos de di- nosaurios hallados en Salitral Moreno (Formación Allen, Cretácico Superior), en la provincia de Río Negro. -

Cairanoolithus: a Large Egg for a Small Dinosaur with Wide Hips
20/11/2020 <em>Cairanoolithus</em>: a Large Egg for a Small Dinosaur With Wide Hips - UABDivulga Barcelona Research & Innovation 02/2015 Cairanoolithus: a Large Egg for a Small Dinosaur With Wide Hips The study of the microstructure of the eggshell of Cairanoolithus conducted by Albert G. Sellés and Angel Galobart, both researchers at the Catalan Institute of Palaeontology Miquel Crusafont (ICP), reveals that this egg type does not belong to sauropod dinosaurs but to ankylosaurs, and probably to Struthiosaurus, a genus of armored dinosaurs. The finding would represent the first description of thyreophora eggs (a group that includes ankylosauria and stegosauria) in the world. It is an arduous task for paleontologists to assign the fossil eggs they found in excavations to a certain dinosaur species or group of dinosaurs, especially because the lack of bones of the parents in the nests and the low probability to find embryonic remains that would help in the taxonomic assignation of the eggs. Thus, we often do not known who laid a certain fossil egg. These limitations have led scientist to design a specific system for naming and classifying fossil eggs, using concepts such as "oogenera" and "ooespecies" for its phylogenetic classification. Cairanoolithus is an oogenus discovered and described in the early 90’s. It takes its name from the first place where it was found, a site near La Cairanne, a small town from southeastern France. From then on, Cairanoolithus has been found in 25 different sites. These eggs are between 72.2 and 71.4 million years old, they are large (over 15 cm in diameter) and have a rounded shape. -

Palaeoecology and Depositional Environments of the Tendaguru Beds (Late Jurassic to Early Cretaceous, Tanzania)
Mitt. Mus. Nat.kd. Berl., Geowiss. Reihe 5 (2002) 19-44 10.11.2002 Palaeoecology and depositional environments of the Tendaguru Beds (Late Jurassic to Early Cretaceous, Tanzania) Martin Aberhan ', Robert Bussert2, Wolf-Dieter Heinrich', Eckhart Schrank2, Stephan Schultkal, Benjamin Sames3, Jiirgen =wet4 & Saidi Kapilima5 With 6 figures, 2 tables, and 2 plates Abstract The Late Jurassic to Early Cretaceous Tendaguru Beds (Tanzania, East Africa) have been well known for nearly a century for their diverse dinosaur assemblages. Here, we present sedimentological and palaeontological data collected by the German- Tanzanian Tendaguru Expedition 2000 in an attempt to reconstruct the palaeo-ecosystems of the Tendaguru Beds at their type locality. Our reconstructions are based on sedimentological data and on a palaeoecological analysis of macroinverte- brates, microvertebrates, plant fossils and microfossils (ostracods, foraminifera, charophytes, palynomorphs). In addition, we included data from previous expeditions, particularly those on the dinosaur assemblages. The environmental model of the Tendaguru Beds presented herein comprises three broad palaeoenvironmental units in a marginal marine setting: (1) Lagoon-like, shallow marine environments above fair weather wave base and with evidence of tides and storms. These formed behind barriers such as ooid bar and siliciclastic sand bar complexes and were generally subject to minor salinity fluctuations. (2) Extended tidal flats and low-relief coastal plains. These include low-energy, brackish coastal lakes and ponds as well as pools and small fluvial channels of coastal plains in which the large dinosaurs were buried. Since these environments apparently were, at best, poorly vegetated, the main feeding grounds of giant sauropods must have been elsewhere. -

Evidence of Reproductive Stress in Titanosaurian
www.nature.com/scientificreports OPEN Evidence of Reproductive Stress in Titanosaurian Sauropods Triggered by an Increase in Ecological Received: 5 April 2017 Accepted: 9 October 2017 Competition Published: xx xx xxxx Albert G. Sellés 1,2, Bernat Vila1,2 & Àngel Galobart1,2 The occurrence of dinosaur pathologic eggs in the Late Cretaceous of Europe is well known, but their origin remains unclear. Here we expose the results of a detailed sampling of the conspicuous fossil record of Late Cretaceous titanosaurian eggs (oogenus Megaloolithius) from several southwestern Europe basins. After examining more than 450 samples, we observed a remarkable and statistically supported occurrence of multiple pathologic eggs in a relatively short stratigraphic range at the end of the early Maastrichtian, circa 71-70 Ma. All pathologic specimens exhibit multi-layered eggshell condition, a characteristic related to dystocia, or egg retention within the female uterus for an abnormal prolonged period of time. After exploring various scenarios, the occurrence of pathologic eggs is strongly correlated with an intense dinosaur faunal replacement that occurred during the early Maastrichtian in the Ibero-Armorican Island. Given that inter-species competiveness is proved to produce major affects in ecological communities, our results suggest that pathologies in the eggs of European titanosaurians could be a consequence of an increase in reproductive stress triggered by direct ecological competition between different dinosaurs. Thus, the present study provides a new perspective of how dinosaurs might have been affected by ecological/environmental disturbance. To understand how species are affected by environmental disturbance is a crucial topic in both modern ecology and palaeoecology. -

Palynology of the Dinosaur Beds of Tendaguru (Tanzania) - Preliminary Results
Mitt. Mus. Nat.kd . Berl., Geowiss. Reihe 2 (1999) 171-183 19.10.1999 Palynology of the Dinosaur Beds of Tendaguru (Tanzania) - Preliminary Results Eckart Schrank' With 2 figures, 3 plates and 2 tables Abstract The Tendaguru Beds, southeastern Tanzania, have yielded two palynological assemblages of Kimmeridgian to Tithonian age: (1) the Anapiculatisporites-Densoisporites-Trisaccites assemblage from the Middle Saurian Beds and (2) the Barbatacysta-Pa- reodinia assemblage from the overlying Smeei Beds. A third assemblage with Rhizophagites and rare angiosperm pollen from the Upper Saurian Beds is contaminated by recent and subrecent material . The Anapiculatisporites-Densoisporites-Trisaccites assemblage is characterized by the presence of freshwater algae (Ovoidi- tes), pteridopyhtic-bryophytic spores and gymnosperm (conifer) pollen, with Classopollis as the most abundant element . Among the rare elements of this assemblage is the questionable dinoflagellate Mendicodinium ? quadratum, possibly a Kim- meridgian-Tithonian marker. The miospores show palaeobiogeographic links to Southern Gondwana, especially Madagascar, Australia, Argentina and India. Deposition of this assemblage took place in an aquatic environment with strong palynological influx from a terrestrial source and questionable marine influence. The Barbatacysta-Pareodinia assemblage contains a considerable number of dinoflagellates suggesting deposition in a mari- ne environment. The terrestrially-derived miospores are impoverished and dominated by conifer pollen, while pteridophytic- bryophytic spores form a very subordinate element or are absent. Key words: Dinoflagellates, pollen, spores, Late Jurassic, Tanzania. Zusammenfassung Die Tendaguru-Schichten, Südost-Tansania, haben zwei palynologische Assoziationen, deren Alter als Kimmeridge bis Tithon interpretiert wird, geliefert . Die Anapiculatisporites-Densoisporites-Trisaccites-Assoziation stammt aus den Mittleren Saurier- schichten, und die Barbatacysta-Pareodinia-Assoziation charakterisiert die darüberlagernden Smeei-Schichten . -

Paleontological Discoveries in the Chorrillo Formation (Upper Campanian-Lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina
Rev. Mus. Argentino Cienc. Nat., n.s. 21(2): 217-293, 2019 ISSN 1514-5158 (impresa) ISSN 1853-0400 (en línea) Paleontological discoveries in the Chorrillo Formation (upper Campanian-lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina Fernando. E. NOVAS1,2, Federico. L. AGNOLIN1,2,3, Sebastián ROZADILLA1,2, Alexis M. ARANCIAGA-ROLANDO1,2, Federico BRISSON-EGLI1,2, Matias J. MOTTA1,2, Mauricio CERRONI1,2, Martín D. EZCURRA2,5, Agustín G. MARTINELLI2,5, Julia S. D´ANGELO1,2, Gerardo ALVAREZ-HERRERA1, Adriel R. GENTIL1,2, Sergio BOGAN3, Nicolás R. CHIMENTO1,2, Jordi A. GARCÍA-MARSÀ1,2, Gastón LO COCO1,2, Sergio E. MIQUEL2,4, Fátima F. BRITO4, Ezequiel I. VERA2,6, 7, Valeria S. PEREZ LOINAZE2,6 , Mariela S. FERNÁNDEZ8 & Leonardo SALGADO2,9 1 Laboratorio de Anatomía Comparada y Evolución de los Vertebrados. Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina - fernovas@yahoo. com.ar. 2 Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina. 3 Fundación de Historia Natural “Felix de Azara”, Universidad Maimonides, Hidalgo 775, C1405BDB Buenos Aires, Argentina. 4 Laboratorio de Malacología terrestre. División Invertebrados Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina. 5 Sección Paleontología de Vertebrados. Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina. 6 División Paleobotánica. Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Avenida Ángel Gallardo 470, Buenos Aires C1405DJR, Argentina. 7 Área de Paleontología. Departamento de Geología, Universidad de Buenos Aires, Pabellón 2, Ciudad Universitaria (C1428EGA) Buenos Aires, Argentina. 8 Instituto de Investigaciones en Biodiversidad y Medioambiente (CONICET-INIBIOMA), Quintral 1250, 8400 San Carlos de Bariloche, Río Negro, Argentina. -
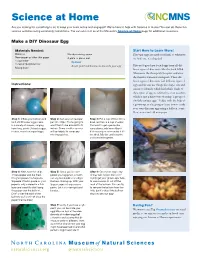
Make a DIY Dinosaur Egg (PDF)
Science at Home Are you looking for something to do to keep your brain active and engaged? We’re here to help with Science at Home! You can do these fun science activities using commonly found items. You can also visit us at the Museum’s Science at Home page for additional resources. Make a DIY Dinosaur Egg Materials Needed: Start Here to Learn More! Balloons Wooden mixing spoon Dinosaur eggs are rarely fossilized, so whenever Newspaper or other thin paper A plate or place-mat we find one, it’s a big deal. 1 cup water Optional 1 cup all-purpose flour Acrylic paint and brushes to decorate your egg Paleontologists have found eggs from all dif- Mixing bowl ferent types of dinosaurs, like the duck-billed Maiasaura, the theropod Oviraptor, and even the massive titanosaur sauropods. These dif- ferent types of dinosaurs laid different types of Instructions: eggs and we can use things like shape, size and texture to identify which laid which. Each of these types of egg are referred to as an ootaxon, which is just a fancy way of saying “a group of similarly unique eggs.” Today, with the help of a grown-up, you’re going to learn how to make your own dinosaur eggs using a balloon, some flour, water and old newspaper. Step 1: Inflate your balloon and Step 2: Cut up your newspa- Step 3: Put a cup of flour into a tie it off. Dinosaur eggs came per into strips. You’re going to bowl, and mix in a cup of water. -

The Tendaguru Formation (Late Jurassic to Early Cretaceous, Southern Tanzania): Definition, Palaeoenvironments, and Sequence Stratigraphy
Fossil Record 12 (2) 2009, 141–174 / DOI 10.1002/mmng.200900004 The Tendaguru Formation (Late Jurassic to Early Cretaceous, southern Tanzania): definition, palaeoenvironments, and sequence stratigraphy Robert Bussert1, Wolf-Dieter Heinrich2 and Martin Aberhan*,2 1 Institut fr Angewandte Geowissenschaften, Technische Universitt Berlin, Skr. BH 2, Ernst-Reuter-Platz 1, 10587 Berlin, Germany. E-mail: [email protected] 2 Museum fr Naturkunde – Leibniz Institute for Research on Evolution and Biodiversity at the Humboldt University Berlin, Invalidenstr. 43, 10115 Berlin, Germany. E-mail: [email protected]; [email protected] Abstract Received 8 December 2008 The well-known Late Jurassic to Early Cretaceous Tendaguru Beds of southern Tanza- Accepted 15 February 2009 nia have yielded fossil plant remains, invertebrates and vertebrates, notably dinosaurs, Published 3 August 2009 of exceptional scientific importance. Based on data of the German-Tanzanian Tenda- guru Expedition 2000 and previous studies, and in accordance with the international stratigraphic guide, we raise the Tendaguru Beds to formational rank and recognise six members (from bottom to top): Lower Dinosaur Member, Nerinella Member, Middle Dinosaur Member, Indotrigonia africana Member, Upper Dinosaur Member, and Ruti- trigonia bornhardti-schwarzi Member. We characterise and discuss each member in de- tail in terms of derivation of name, definition of a type section, distribution, thickness, lithofacies, boundaries, palaeontology, and age. The age of the whole formation appar- ently ranges at least from the middle Oxfordian to the Valanginian through Hauterivian or possibly Aptian. The Tendaguru Formation constitutes a cyclic sedimentary succes- sion, consisting of three marginal marine, sandstone-dominated depositional units and three predominantly coastal to tidal plain, fine-grained depositional units with dinosaur remains. -

The First Dinosaur Egg Remains a Mystery
bioRxiv preprint doi: https://doi.org/10.1101/2020.12.10.406678; this version posted December 11, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 The first dinosaur egg remains a mystery 2 3 Lucas J. Legendre1*, David Rubilar-Rogers2, Alexander O. Vargas3, and Julia A. 4 Clarke1* 5 6 1Department of Geological Sciences, University of Texas at Austin, Austin, Texas 78756, 7 USA. 8 2Área Paleontología, Museo Nacional de Historia Natural, Casilla 787, Santiago, Chile. 9 3Departamento de Biología, Facultad de Ciencias, Universidad de Chile, Santiago 7800003, 10 Chile. 11 1 bioRxiv preprint doi: https://doi.org/10.1101/2020.12.10.406678; this version posted December 11, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 12 Abstract 13 A recent study by Norell et al. (2020) described new egg specimens for two dinosaur species, 14 identified as the first soft-shelled dinosaur eggs. The authors used phylogenetic comparative 15 methods to reconstruct eggshell type in a sample of reptiles, and identified the eggs of 16 dinosaurs and archosaurs as ancestrally soft-shelled, with three independent acquisitions of a 17 hard eggshell among dinosaurs. This result contradicts previous hypotheses of hard-shelled 18 eggs as ancestral to archosaurs and dinosaurs. -
![[PDF] Sedimentology and Depositional Environments of the Red](https://docslib.b-cdn.net/cover/3942/pdf-sedimentology-and-depositional-environments-of-the-red-1553942.webp)
[PDF] Sedimentology and Depositional Environments of the Red
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright Author's personal copy Journal of African Earth Sciences 57 (2010) 179–212 Contents lists available at ScienceDirect Journal of African Earth Sciences journal homepage: www.elsevier.com/locate/jafrearsci Geological Society of Africa Presidential Review No. 15 Sedimentology and depositional environments of the Red Sandstone Group, Rukwa Rift Basin, southwestern Tanzania: New insight into Cretaceous and Paleogene terrestrial ecosystems and tectonics in sub-equatorial Africa Eric M. Roberts a,*, Patrick M. O’Connor b, Nancy J. Stevens b, Michael D. Gottfried c, Zubair A. Jinnah d, Sifael Ngasala c, Adeline M. Choh d, Richard A. Armstrong e a Department of Physical Sciences, Southern Utah University, Cedar City, UT 84720, USA b Department of Biomedical Sciences, 228 Irvine Hall, College of Osteopathic Medicine, Ohio University, Athens, OH, 45701,