JBES-Vol6no1-P469-47
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Our Brochure
Fifty years ago our National Bird, the Blue Crane, was a common sight in South Africa's grasslands, today it is rare. The KZN Crane Foundation, was formed to understand why and find ways to reverse this trend. inside front cover The KZN Crane Foundation By the year 2000, the number of Blue Cranes in the grasslands of the Eastern Cape, KZN, the Free State, Gauteng, and Mpumalanga, had declined by 90%. Their breeding requirements are, however, flexible and they have been able to adapt to new habitats in the Karoo and the wheat-lands of the Southern Cape and the total national population appears, for now, although greatly reduced, to be stable. Wattled Cranes (our largest and most regal species), unlike Blue Cranes, are much more specific about their nesting requirements (needing wetlands) and have been unable to adapt to other habitats. Like the Blue Cranes their population plummeted during the Nineteen eighties and nineties and by the year 2000, only 80 breeding pairs remained, an almost unsustainable level. At the time, their extinction seemed inevitable. In light of this, in 1989 concerned conservationists, under the leadership of the then Natal Parks Board, formed the KZN Crane Foundation with the aim of understanding and reversing this decline. (Photo Thanks to Daniel Dolpire) Are Cranes Important? Cranes are ancient birds whose elegance and beauty have long captured man's imagination, with references to them in the bible and the mythologies of many cultures, as symbols of peace, fidelity, and longevity. South African legend has it that Shaka wore Blue Crane feathers in his head-dress and decreed that feathers of these magnificent birds may be worn only by kings. -

Onetouch 4.0 Scanned Documents
/ Chapter 2 THE FOSSIL RECORD OF BIRDS Storrs L. Olson Department of Vertebrate Zoology National Museum of Natural History Smithsonian Institution Washington, DC. I. Introduction 80 II. Archaeopteryx 85 III. Early Cretaceous Birds 87 IV. Hesperornithiformes 89 V. Ichthyornithiformes 91 VI. Other Mesozojc Birds 92 VII. Paleognathous Birds 96 A. The Problem of the Origins of Paleognathous Birds 96 B. The Fossil Record of Paleognathous Birds 104 VIII. The "Basal" Land Bird Assemblage 107 A. Opisthocomidae 109 B. Musophagidae 109 C. Cuculidae HO D. Falconidae HI E. Sagittariidae 112 F. Accipitridae 112 G. Pandionidae 114 H. Galliformes 114 1. Family Incertae Sedis Turnicidae 119 J. Columbiformes 119 K. Psittaciforines 120 L. Family Incertae Sedis Zygodactylidae 121 IX. The "Higher" Land Bird Assemblage 122 A. Coliiformes 124 B. Coraciiformes (Including Trogonidae and Galbulae) 124 C. Strigiformes 129 D. Caprimulgiformes 132 E. Apodiformes 134 F. Family Incertae Sedis Trochilidae 135 G. Order Incertae Sedis Bucerotiformes (Including Upupae) 136 H. Piciformes 138 I. Passeriformes 139 X. The Water Bird Assemblage 141 A. Gruiformes 142 B. Family Incertae Sedis Ardeidae 165 79 Avian Biology, Vol. Vlll ISBN 0-12-249408-3 80 STORES L. OLSON C. Family Incertae Sedis Podicipedidae 168 D. Charadriiformes 169 E. Anseriformes 186 F. Ciconiiformes 188 G. Pelecaniformes 192 H. Procellariiformes 208 I. Gaviiformes 212 J. Sphenisciformes 217 XI. Conclusion 217 References 218 I. Introduction Avian paleontology has long been a poor stepsister to its mammalian counterpart, a fact that may be attributed in some measure to an insufRcien- cy of qualified workers and to the absence in birds of heterodont teeth, on which the greater proportion of the fossil record of mammals is founded. -

Wood-Stork-2005.Pdf
WOOD STORK Mycteria americana Photo of adult wood stork in landing posture and chicks in Close-up photo of adult wood stork. nest. Photo courtesy of USFWS/Photo by George Gentry Photo courtesy of U.S. Army Corps of Engineers. FAMILY: Ciconiidae STATUS: Endangered - U.S. Breeding Population (Federal Register, February 28, 1984) [Service Proposed for status upgrade to Threatened, December 26, 2013] DESCRIPTION: Wood storks are large, long-legged wading birds, about 45 inches tall, with a wingspan of 60 to 65 inches. The plumage is white except for black primaries and secondaries and a short black tail. The head and neck are largely unfeathered and dark gray in color. The bill is black, thick at the base, and slightly decurved. Immature birds have dingy gray feathers on their head and a yellowish bill. FEEDING HABITS: Small fish from 1 to 6 inches long, especially topminnows and sunfish, provide this bird's primary diet. Wood storks capture their prey by a specialized technique known as grope-feeding or tacto-location. Feeding often occurs in water 6 to 10 inches deep, where a stork probes with the bill partly open. When a fish touches the bill it quickly snaps shut. The average response time of this reflex is 25 milliseconds, making it one of the fastest reflexes known in vertebrates. Wood storks use thermals to soar as far as 80 miles from nesting to feeding areas. Since thermals do not form in early morning, wood storks may arrive at feeding areas later than other wading bird species such as herons. -

Colonial Nesting Waterbirds Fact Sheet
U.S. Fish & Wildlife Service Colonial-Nesting Waterbirds A Glorious and Gregarious Group What Is a Colonial-Nesting Waterbird? species of cormorants, gulls, and terns also Colonial-nesting waterbird is a tongue- occupy freshwater habitats. twister of a collective term used by bird biologists to refer to a large variety of Wading Birds seek their prey in fresh or different species that share two common brackish waters. As the name implies, characteristics: (1) they tend to gather in these birds feed principally by wading or large assemblages, called colonies, during standing still in the water, patiently waiting the nesting season, and (2) they obtain all for fish or other prey to swim within Migratory Bird Management or most of their food (fish and aquatic striking distance. The wading birds invertebrates) from the water. Colonial- include the bitterns, herons, egrets, night- nesting waterbirds can be further divided herons, ibises, spoonbills, and storks. Mission into two major groups depending on where they feed. Should We Be Concerned About the To conserve migratory bird Conservation Status of Colonial-Nesting populations and their habitats Seabirds (also called marine birds, oceanic Waterbirds? birds, or pelagic birds) feed primarily in Yes. While many species of seabirds for future generations, through saltwater. Some seabirds are so appear to have incredibly large careful monitoring and effective marvelously adapted to marine populations, they nevertheless face a management. environments that they spend virtually steady barrage of threats, such as oil their entire lives at sea, returning to land pollution associated with increased tanker only to nest; others (especially the gulls traffic and spills, direct mortality from and terns) are confined to the narrow entanglement and drowning in commercial coastal interface between land and sea, fishing gear, depletion of forage fishes due feeding during the day and loafing and to overexploitation by commercial roosting on land. -

Spread-Wing Postures and Their Possible Functions in the Ciconiidae
THE AUK A QUARTERLY JOURNAL OF ORNITHOLOGY Von. 88 Oc:roBE'a 1971 No. 4 SPREAD-WING POSTURES AND THEIR POSSIBLE FUNCTIONS IN THE CICONIIDAE M. P. KAI-IL IN two recent papers Clark (19'69) and Curry-Lindahl (1970) have reported spread-wingpostures in storks and other birds and discussed someof the functionsthat they may serve. During recent field studies (1959-69) of all 17 speciesof storks, I have had opportunitiesto observespread-wing postures. in a number of speciesand under different environmentalconditions (Table i). The contextsin which thesepostures occur shed somelight on their possible functions. TYPES OF SPREAD-WING POSTURES Varying degreesof wing spreadingare shownby at least 13 species of storksunder different conditions.In somestorks (e.g. Ciconia nigra, Euxenuragaleata, Ephippiorhynchus senegalensis, and ]abiru mycteria) I observedno spread-wingpostures and have foundno referenceto them in the literature. In the White Stork (Ciconia ciconia) I observedonly a wing-droopingposture--with the wings held a short distanceaway from the sidesand the primaries fanned downward--in migrant birds wetted by a heavy rain at NgorongoroCrater, Tanzania. Other species often openedthe wingsonly part way, in a delta-wingposture (Frontis- piece), in which the forearmsare openedbut the primariesremain folded so that their tips crossin front o.f or below the. tail. In some species (e.g. Ibis leucocephalus)this was the most commonly observedspread- wing posture. All those specieslisted in Table i, with the exception of C. ciconia,at times adopted a full-spreadposture (Figures i, 2, 3), similar to those referred to by Clark (1969) and Curry-Lindahl (1970) in severalgroups of water birds. -

Conservation Measures for the Siberian Crane
CMS Technical Series Publication No. 1 Conservation Measures for the Siberian Crane Convention on Migratory Species Published by: UNEP/CMS Secretariat, Bonn, Germany Recommended citation: UNEP/CMS. ed.(1999). Conservation Measures for the Siberian Crane. CMS Technical Series Publication No.1, UNEP/CMS Secretariat, Bonn, Germany. Cover photograph: Siberian Crane (Grus leucogeranus) in snow. © Sietre / BIOS, Paris © UNEP/CMS, 1999 (copyright of individual contributions remains with the authors). Reproduction of this publication, except the cover photograph, for educational and other non-commercial purposes is authorized without permission from the copyright holder, provided the source is cited and the copyright holder receives a copy of the reproduced material. Reproduction of the text for resale or other commercial purposes, or of the cover photograph, is prohibited without prior permission of the copyright holder. The views expressed in this publication are those of the authors and do not necessarily reflect the views or policies of UNEP/CMS, nor are they an official record. The designation of geographical entities in this publication, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of UNEP/CMS concerning the legal status of any country, territory or area, or of its authorities, nor concerning the delimitation of its frontiers and boundaries. Copies of this publication are available from the UNEP/CMS Secretariat, United Nations Premises in Bonn, Martin-Luther-King-Str. 8, D-53175 -

Imperiled Coastal Birds of Florida and the State Laws That Protect Them
Reddish Egret Roseate Spoonbill Threatened (S) Threatened (S) Imperiled Coastal The rarest heron in North Using spatulala-shaped Birds of Florida America, Reddish Egrets bills to feel prey in shallow are strictly coastal. They ponds, streams, or coastal and the chase small fish on open waters, Roseate Spoonbills State Laws that flats. They nest in small nest in trees along the numbers on estuary coast and inland. Having Protect Them islands, usually in colonies barely recovered from with other nesting wading hunting eradication, these birds. This mid-sized heron birds now face extirpation is mostly gray with rust- from climate change and colored head, though some sea-level rise. birds are solid white. Wood Stork Florida Sandhill Threatened (F) Florida Statutes and Rules Crane This large wading bird Threatened (S) is the only stork in the 68A-27.003 Designation and management of the state- This crane subspecies is Americas. Breeding areas listed species and coordination with federal government for resident year-round in have shifted from south federally-listed species Florida, and defends a Florida and the Everglades nesting territory that is northward. Wood Storks 68A-19.005 General Regulations relating to state- must have abundant prey adjacent to open upland designated Critical Wildlife Areas foraging habitat. Nesting concentrated in shallow in shallow ponds, adults wetlands in order to feed 68A-4.001 Controls harvest of wildlife only under permitted defend their eggs or chicks their young. Prey items from predators including include -

Birds (DNA'dna Hybridization/Mtdna Sequences/Phylogeny/Systematics)
Proc. Natl. Acad. Sci. USA Vol. 91, pp. 9861-9865, October 1994 Evolution Molecules vs. morphology in avian evolution: The case of the "pelecaniform" birds (DNA'DNA hybridization/mtDNA sequences/phylogeny/systematics) S. BLAIR HEDGES* AND CHARLES G. SIBLEyt *Department of Biology and Institute of Molecular Evolutionary Genetics, 208 Mueller Laboratory, Pennsylvania State University, University Park, PA 16802; and t433 Woodley Place, Santa Rosa, CA 95409 Contributed by Charles G. Sibley, June 20, 1994 ABSTRACT The traditional avian Order Pelecaniformes the three front toes has evolved in groups with separate is composed of birds with all four toes connected by a web. This origins-e.g., ducks, gulls, flamingos, and albatrosses. Could "totipalmate" condition is found in ca. 66 living species: 8 the totipalmate condition, which occurs in fewer species, also pelicans (Pelecanus), 9 boobies and gannets (Sula, Papasula, have multiple origins? Sibley and Ahlquist (2) reviewed the Morus), ca. 37 cormorants (Phalacrocorax) , 4 anhingas or literature from 1758 to 1990. darters (Anhinga), 5 frigatebirds (Fregata), and 3 tropicbirds There have been many morphological studies of the pele (Phaethon). Several additional characters are shared by these caniforms; those of Lanham (3), Saiff(4), and Cracraft (5) are genera, and their monophyly has been assumed since the among the most recent. Lanham (3) recognized their diversity beginning of modern zoological nomenclature. Most ornithol but concluded that the totipalmate birds form a natural order. ogists classify these genera as an order, although tropicbirds He assigned Phaethon and Fregata to separate suborders, have been viewed as related to terns, and frigatebirds as the other genera to the suborder Pelecani, and suggested that relatives of the petrels and albatrosses. -

Alpha Codes for 2168 Bird Species (And 113 Non-Species Taxa) in Accordance with the 62Nd AOU Supplement (2021), Sorted Taxonomically
Four-letter (English Name) and Six-letter (Scientific Name) Alpha Codes for 2168 Bird Species (and 113 Non-Species Taxa) in accordance with the 62nd AOU Supplement (2021), sorted taxonomically Prepared by Peter Pyle and David F. DeSante The Institute for Bird Populations www.birdpop.org ENGLISH NAME 4-LETTER CODE SCIENTIFIC NAME 6-LETTER CODE Highland Tinamou HITI Nothocercus bonapartei NOTBON Great Tinamou GRTI Tinamus major TINMAJ Little Tinamou LITI Crypturellus soui CRYSOU Thicket Tinamou THTI Crypturellus cinnamomeus CRYCIN Slaty-breasted Tinamou SBTI Crypturellus boucardi CRYBOU Choco Tinamou CHTI Crypturellus kerriae CRYKER White-faced Whistling-Duck WFWD Dendrocygna viduata DENVID Black-bellied Whistling-Duck BBWD Dendrocygna autumnalis DENAUT West Indian Whistling-Duck WIWD Dendrocygna arborea DENARB Fulvous Whistling-Duck FUWD Dendrocygna bicolor DENBIC Emperor Goose EMGO Anser canagicus ANSCAN Snow Goose SNGO Anser caerulescens ANSCAE + Lesser Snow Goose White-morph LSGW Anser caerulescens caerulescens ANSCCA + Lesser Snow Goose Intermediate-morph LSGI Anser caerulescens caerulescens ANSCCA + Lesser Snow Goose Blue-morph LSGB Anser caerulescens caerulescens ANSCCA + Greater Snow Goose White-morph GSGW Anser caerulescens atlantica ANSCAT + Greater Snow Goose Intermediate-morph GSGI Anser caerulescens atlantica ANSCAT + Greater Snow Goose Blue-morph GSGB Anser caerulescens atlantica ANSCAT + Snow X Ross's Goose Hybrid SRGH Anser caerulescens x rossii ANSCAR + Snow/Ross's Goose SRGO Anser caerulescens/rossii ANSCRO Ross's Goose -

The Flight of Albatross—How to Transform It Into Aerodynamic Engineering?
Engineering, 2014, 6, 427-438 Published Online July 2014 in SciRes. http://www.scirp.org/journal/eng http://dx.doi.org/10.4236/eng.2014.68045 The Flight of Albatross—How to Transform It into Aerodynamic Engineering? Günther Pfeifhofer*, Helmut Tributsch Carinthia University of Applied Sciences, Villach, Austria Email: *[email protected], [email protected] Received 26 April 2014; revised 30 May 2014; accepted 12 June 2014 Copyright © 2014 by authors and Scientific Research Publishing Inc. This work is licensed under the Creative Commons Attribution International License (CC BY). http://creativecommons.org/licenses/by/4.0/ Abstract The flight of albatross (Diomedea exulans) takes advantage of the up-drift which is determined by the product of relative wind velocity and it’s gradient above the sea surface, to power its elegant (dynamic) flight over the ocean. Some of the complicated flight manoeuvres are determined by biological necessities. From its most basic flight manoeuvre a technical aerodynamically scheme can be derived which allows the design of a mechanical technical prototype of a wind generator. It is based on a rotational movement in combination with a skillful time dependent adjustment of the airfoil. Several technical possibilities are discussed and with one of these elaborated in some detail. The technology to be developed could be applied in highly asymmetric air streaming envi- ronment around high rise buildings, on mountain ridges and of course, also low above sea level and plains. Mathematical-technical conditions for power gain are discussed. The technology could, in principle, also be deployed to exploit velocity gradients in river water environment. -
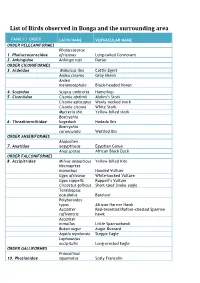
Birds Species in Bonga List
List of Birds observed in Bonga and the surrounding area FAMILY / ORDER LATIN NAME VERNACULAR NAME ORDER PELECANIFORMES Phalacrocorax 1. Phalacrocoracidae africanus Long-tailed Cormorant 2. Anhingidae Anhinga rufa Darter ORDER CICONIIFORMES 3. Ardeidae Bubulcus ibis Cattle Egret Ardea cinerea Grey Heron Ardea melanocephala Black-headed Heron 4. Scopidae Scopus umbretta Hamerkop 5. Ciconiidae Ciconia abdimii Abdim’s Stork Ciconia episcopus Wooly -necked stork Ciconia ciconia White Stork Mycteria ibis Yellow -billed stork Bostrychia 6. Threskiornithidae hagedash Hadada Ibis Bostrychia carunculata Wattled Ibis ORDER ANSERIFORMES Alopochen 7. Anatidae aegyptiacus Egyptian Goose Anas sparsa African Black Duck ORDER FALCONIFORMES 8. Accipitridae Milvus aegypticus Yellow -billed Kite Necrosyrtes monachus Hooded Vulture Gyps africanus White -backed Vulture Gyps ruppellii Ruppell’s Vulture Circaetus gallicus Short -toed Snake -eagle Terathopius ecaudatus Bateleur Polyboroides typus African Harrier Hawk Accipiter Red -breasted/Rufous -chested Sparrow rufiventris hawk Accipiter minullus Little Sparrowhawk Buteo augur Augur Buzzard Aquila nipalensis Steppe Eagle Lophoaetus occipitalis Long-crested Eagle ORDER GALLIFORMES Francolinus 10. Phasianidae squamatus Scaly Francolin FAMILY / ORDER LATIN NAME VERNACULAR NAME Francolinus castaneicollis Chestnut-napped Francolin ORDER GRUIFORMES Balearica 11. Gruidae pavonina Black Crowned Crane Bugeranus carunculatus Wattled crane Ruogetius 12. Rallidae rougetii Rouget’s Rail Podica 13. Heliornithidae senegalensis African Finfoot ORDER CHARADRIIFORMES 14. Scolopacidae Tringa ochropus Green Sandpiper Tringa hypolucos Common Sandpiper ORDER COLUMBIFORMES 15. Columbidae Columba guinea Speckled Pigeon Columba arquatrix African Olive Pigeon (Rameron Pigeon) Streptopelia lugens Dusky (Pink-breasted) Turtle Dove Streptopelia semitorquata Red-eyed Dove Turtur chalcospilos Emerald-spotted Wood Dove Turtur tympanistria Tambourine Dove ORDER CUCULIFORMES 17. Musophagidae Tauraco leucotis White -cheeked Turaco Centropus 18. -

2020 National Bird List
2020 NATIONAL BIRD LIST See General Rules, Eye Protection & other Policies on www.soinc.org as they apply to every event. Kingdom – ANIMALIA Great Blue Heron Ardea herodias ORDER: Charadriiformes Phylum – CHORDATA Snowy Egret Egretta thula Lapwings and Plovers (Charadriidae) Green Heron American Golden-Plover Subphylum – VERTEBRATA Black-crowned Night-heron Killdeer Charadrius vociferus Class - AVES Ibises and Spoonbills Oystercatchers (Haematopodidae) Family Group (Family Name) (Threskiornithidae) American Oystercatcher Common Name [Scientifc name Roseate Spoonbill Platalea ajaja Stilts and Avocets (Recurvirostridae) is in italics] Black-necked Stilt ORDER: Anseriformes ORDER: Suliformes American Avocet Recurvirostra Ducks, Geese, and Swans (Anatidae) Cormorants (Phalacrocoracidae) americana Black-bellied Whistling-duck Double-crested Cormorant Sandpipers, Phalaropes, and Allies Snow Goose Phalacrocorax auritus (Scolopacidae) Canada Goose Branta canadensis Darters (Anhingidae) Spotted Sandpiper Trumpeter Swan Anhinga Anhinga anhinga Ruddy Turnstone Wood Duck Aix sponsa Frigatebirds (Fregatidae) Dunlin Calidris alpina Mallard Anas platyrhynchos Magnifcent Frigatebird Wilson’s Snipe Northern Shoveler American Woodcock Scolopax minor Green-winged Teal ORDER: Ciconiiformes Gulls, Terns, and Skimmers (Laridae) Canvasback Deep-water Waders (Ciconiidae) Laughing Gull Hooded Merganser Wood Stork Ring-billed Gull Herring Gull Larus argentatus ORDER: Galliformes ORDER: Falconiformes Least Tern Sternula antillarum Partridges, Grouse, Turkeys, and