Taxonomic and Ecologic Properties of the Endangered Iris Pamphylica (Iridaceae) Endemic to S.W. Anatolia INTRODUCTION MATERIALS
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
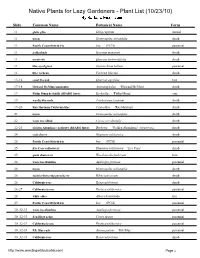
Native Plants for Lazy Gardeners - Plant List (10/23/10)
Native Plants for Lazy Gardeners - Plant List (10/23/10) Slide Common Name Botanical Name Form 11 globe gilia Gilia capitata annual 11 toyon Heteromeles arbutifolia shrub 11 Pacific Coast Hybrid iris Iris (PCH) perennial 11 goldenbush Isocoma menziesii shrub 11 scrub oak Quercus berberidifolia shrub 11 blue-eyed grass Sisyrinchium bellum perennial 11 lilac verbena Verbena lilacina shrub 13-16 coast live oak Quercus agrifolia tree 17-18 Howard McMinn man anita Arctostaphylos 'Howard McMinn' shrub 19 Philip Mun keckiella (RSABG Intro) Keckiella 'Philip Munz' ine 19 woolly bluecurls Trichostema lanatum shrub 19-20 Ray Hartman California lilac Ceanothus 'Ray Hartman' shrub 21 toyon Heteromeles arbutifolia shrub 22 western redbud Cercis occidentalis shrub 22-23 Golden Abundance barberry (RSABG Intro) Berberis 'Golden Abundance' (MAHONIA) shrub 2, coffeeberry Rhamnus californica shrub 25 Pacific Coast Hybrid iris Iris (PCH) perennial 25 Eve Case coffeeberry Rhamnus californica '. e Case' shrub 25 giant chain fern Woodwardia fimbriata fern 26 western columbine Aquilegia formosa perennial 26 toyon Heteromeles arbutifolia shrub 26 fuchsia-flowering gooseberry Ribes speciosum shrub 26 California rose Rosa californica shrub 26-27 California fescue Festuca californica perennial 28 white alder Alnus rhombifolia tree 29 Pacific Coast Hybrid iris Iris (PCH) perennial 30 032-33 western columbine Aquilegia formosa perennial 30 032-33 San Diego sedge Carex spissa perennial 30 032-33 California fescue Festuca californica perennial 30 032-33 Elk Blue rush Juncus patens '.l1 2lue' perennial 30 032-33 California rose Rosa californica shrub http://www weedingwildsuburbia com/ Page 1 30 032-3, toyon Heteromeles arbutifolia shrub 30 032-3, fuchsia-flowering gooseberry Ribes speciosum shrub 30 032-3, Claremont pink-flowering currant (RSA Intro) Ribes sanguineum ar. -

A HANDBOOK of GARDEN IRISES by W
A HANDBOOK OF GARDEN IRISES By W. R. DYKES, M.A., L.-ès-L. SECRETARY OF THE ROYAL HORTICULTURAL SOCIETY. AUTHOR OF "THE GENUS IRIS," ETC. CONTENTS. PAGE PREFACE 3 1 THE PARTS OF THE IRTS FLOWER AND PLANT 4 2 THE VARIOUS SECTIONS OF THE GENUS AND 5 THEIR DISTRIBUTION 3 THE GEOGRAPHICAL DISTRIBUTION OF THE VARIOUS 10 SECTIONS AND SPECIES AND THEIR RELATIVE AGES 4 THE NEPALENSIS SECTION 13 5 THE GYNANDRIRIS SECTION 15 6 THE RETICULATA SECTION 16 7 THE JUNO SECTION 23 8 THE XIPHIUM SECTION 33 9 THE EVANSIA SECTION 40 10 THE PARDANTHOPSIS SECTION 45 11 THE APOGON SECTION 46 — THE SIBIRICA SUBSECTION 47 — THE SPURIA SUBSECTION 53 — THE CALIFORNIAN SUBSECTION 59 — THE LONGIPETALA SUBSECTION 64 — THE HEXAGONA SUBSECTION 67 — MISCELLANEOUS BEARDLESS IRISES 69 12 THE ONCOCYCLUS SECTION 77 I. Polyhymnia, a Regeliocydus hybrid. 13 THE REGELIA SECTION 83 (I. Korolkowi x I. susianna). 14 THE PSEUDOREGELIA SECTION 88 15 THE POGONIRIS SECTION 90 16 GARDEN BEARDED IRISES 108 17 A NOTE ON CULTIVATION, ON RAISING 114 SEEDLINGS AND ON DISEASES 18 A TABLE OF TIMES OF PLANTING AND FLOWERING 116 19 A LIST OF SYNONYMS SOMETIMES USED IN 121 GARDENS This edition is copyright © The Goup for Beardless Irises 2009 - All Rights Reserved It may be distributed for educational purposes in this format as long as no fee (or other consideration) is involved. www.beardlessiris.org PREFACE TO THIS DIGITAL EDITION William Rickatson Dykes (1877-1925) had the advantage of growing irises for many years before writing about them. This Handbook published in 1924 represents the accumulation of a lifetime’s knowledge. -

Philip J. Savage, Jr
ISSUE 73 M4SNOU4 Philip J. Savage, Jr. May 8, 1917 — October 13, 2002 Phil Savnge, /r. , an icon m the Magnolia worhl, passed au ay last October of conr plica- lions from the West Nile virus. Phil, a renotvrred magnolin Irybcidrzer, specialized in breeding mngnolias that u&cre cold Irardy. Becnuse of this, nragnolia entirusiasts living in colder climalcs now have morry rrrorc choices thnn did Dennis Ledvh rn u&hen he bought Iris lrouse irr Green Bay, Wiscorrsin in thc Intr typos Plril hns left an errduring legaclt n&itlr tire mnny /Inc, cold-lurrdy hybrids Ire bred. Following arc set&eral rwni niscclrces fronr irrdividuals who toere deqviy in/luencerl by Phil mrd Iris u&ork u&ith mngrrolins. DENNIs LEDVINA WIUTES. Back in the late yos the landscaping around my new house consisted of three magnolias: two M. x sorrlarrgeana, and a M. x loebneri 'MerrilL' At the time, these were the only magnolias gener- ally available at local nurseries. As I watched these magnolias bloom each spring, I became more intrigued with their beautiful Bowers and began driv- ing around Green Bay to observe and admire some of the established trees. My admiration for the genus contin- ued to grow each year as I began col- lecting more information about these magnificent plants. One summer I was in the Detroit area and I decided to call this magnolia expert, Phil Savage, that I had read so much about. I can vividly remember calling Phil from a telephone booth on Telegraph Road and finding mysell, an unknown amateur, talking to a magnolia expert who from the first made me feel like a lifelong friend. -

AGS Seed List No 69 2020
Seed list No 69 2020-21 Garden Collected Seed 1001 Abelia floribunda 1057 Agrostemma githago 1002 Abies koreana 1058 Albuca canadensis (L. -

May 15, 2016 Passing Peony and Iris Plants on from Generation to Generation Annette Meyer Heisdorffer Daviess County Extension Agent for Horticulture
May 15, 2016 Passing Peony and Iris Plants on from Generation to Generation Annette Meyer Heisdorffer Daviess County Extension Agent for Horticulture After lunch on Mother’s Day, my mom and I surveyed her garden, especially the peonies. We both agreed that I needed to propagate her peonies and plant them in my garden. These are special, because I remember them growing in my grandmother’s garden. Peonies are commonly passed down from generation to generation. My goal is to someday share them with my twins. Our discussion included the irises, which are another heritage plant. Both plants are blooming beautifully in May and are spectacular in the garden. Information about these two plants will be provided in this article. Peony (Paeonia officinalis, Paeonia lactiflora, and hybrids) is a herbaceous perennial, which means at the end of the growing season it will die back to the ground. However, the plant returns year after year. Peonies grow best in full sun and well-drained soil. There are tree peonies (Paeonia suffruticosa) which have a woody stem, but those are not as common and require different growing conditions. The tree peony will not be discussed here. According to Dr. Rick Durham, Extension Specialist for Consumer Horticulture, peonies can be found in landscapes across Kentucky. Peonies have a long life span and are commonly grown in the garden. When planting the root, make sure it is not too deep. The eyes or bud should be just below the surface of the soil. If it is planted too deeply, the plants won't bloom. -

Cappadocia Wildlife Tour Report Turkey Botanical Birdwatching
Hatay to Cappadocia Flowers, Birds & Ancient Sites of Turkey A Greentours Trip Report 12th to 23rd May 2012 Led by Seda Soylu and Phil Benstead The following report is from the itinerary we did in 2012. The 2013 tour differs in that instead of visiting the Hatay region in 2013 we’ll be starting the tour in Konya and Kulu Golu in the centre of Anatolia and the rest of the tour will be done in reverse of the 2012 order. So the most relevant days are days 5-12 in this report, and these correspond to days 4-11 (but in reverse order!) in the 2013 itinerary. Daily report written by Phil Benstead and Seda Soylu Day 1 Saturday 12th May arrival We all converged on the airport at Istanbul by different routes, negotiating the visa buying process and the march across to the domestic terminal. The deciding game between Galatasaray and Fenerbahçe in the Turkish Championship took place in Istanbul during the evening and even the airport departure area was caught up in the excitement. Very atmospheric. We eventually all met up at the gate for our late night onward flight and then dozed through the short journey to Hatay and were soon being met by a smiling Seda and whisked off in the minibus to the hotel. Day 2 Sunday 13th May the road to Yayladağı A leisurely start to the day was required this morning after the late (early?) arrival at the hotel last night. Breakfasted we headed for the van and drove south towards Syria on the road to Yayladağ ı. -

Basic Guide to the Culture of Peonies This Is Easily Done by Pulling Them Downwards and Sideways with the Fingers
A handful of sheep manure to a plant may be given in the spring to improve the bloom. Liquid manure also may be used with discretion, for the same purpose. Disbudding. Most varieties of peonies develop several small lateral or America’s Largest Grower of Daylilies, Peonies and Iris sidebuds near the base of the terminal bud. If large flowers are wanted, the side buds should be removed so the strength will all go into the terminal bud. The side-buds should be removed as soon as they are about the size of a pea. Basic Guide to the Culture of Peonies This is easily done by pulling them downwards and sideways with the fingers. o Some people prefer leaving their side-buds which develop and prolong the P.O. Box 338 • Sarcoxie, MO 64862 • (417) 548-3514 blooming season. The side-buds bloom later than the terminal buds. Insect Pests. In some sections of the country, where thrips are Basic Guide to the Culture of Peonies prevalent, some late varieties are damaged to the extent that the buds fail to open even after they are almost fully developed. Spraying or dusting, at Peonies are easily grown and their requirements are few, but they respond weekly intervals should control the thrips very well. Apply first application beautifully to a little special care and attention by producing best quality when buds are about the size of a large marble. We like Orthene. flowers and many of them. With this thought in mind we offer the following suggestions gained from many years experience in growing and Planted too deeply...examine and if Why Peonies Do Not Bloom. -

Document Converted With
Southwest Turkey A Greentours Trip Report 21st March – 2nd April 2010 Led by Başak Gardner Day 1 Journey to Antalya 21.03.10 I met the group at the airport and directly drove to the hotel. It was around 10 pm. Day 2 Journey to Ibradı 22.03.10 With good news from the meteorology the tour started. Alpine Swifts were flying around and Yellow-vented Bulbuls were singing from the roof as we were having our breakfast by the pool. A very short visit to the bank to change and get some money gave a chance for the group to take some pictures of the castle walls and gate. We even did some botanizing. The walls of the very old houses in the old town were almost covered by Cymbalaria longipes. We were on the road before 10 am and our first stop was by a graveyard where Pyrus serikensis, which is an endangered endemic pear tree, was in full bloom. But the main plant we were looking for was a bit over, however we managed to find some fresh reticulately-patterned Iris masia. Along with these were some Muscari comosum, Anemone coronaria and a single plant of Gladiolus italicus. Val spotted our first butterfly an Eastern Festoon and both Large White and Orange Tip were also seen as well as Danford’s Lizard. We made another stop both to have lunch and to look for some Ophrys. Along an old track by the picnic site we encountered many Ophrys mammosa spikes in good flower. We had a good lunch with some Turkish tea our driver prepared for us among the Bellis annua flowers. -

These De Doctorat De L'universite Paris-Saclay
NNT : 2016SACLS250 THESE DE DOCTORAT DE L’UNIVERSITE PARIS-SACLAY, préparée à l’Université Paris-Sud ÉCOLE DOCTORALE N° 567 Sciences du Végétal : du Gène à l’Ecosystème Spécialité de doctorat (Biologie) Par Mlle Nour Abdel Samad Titre de la thèse (CARACTERISATION GENETIQUE DU GENRE IRIS EVOLUANT DANS LA MEDITERRANEE ORIENTALE) Thèse présentée et soutenue à « Beyrouth », le « 21/09/2016 » : Composition du Jury : M., Tohmé, Georges CNRS (Liban) Président Mme, Garnatje, Teresa Institut Botànic de Barcelona (Espagne) Rapporteur M., Bacchetta, Gianluigi Università degli Studi di Cagliari (Italie) Rapporteur Mme, Nadot, Sophie Université Paris-Sud (France) Examinateur Mlle, El Chamy, Laure Université Saint-Joseph (Liban) Examinateur Mme, Siljak-Yakovlev, Sonja Université Paris-Sud (France) Directeur de thèse Mme, Bou Dagher-Kharrat, Magda Université Saint-Joseph (Liban) Co-directeur de thèse UNIVERSITE SAINT-JOSEPH FACULTE DES SCIENCES THESE DE DOCTORAT DISCIPLINE : Sciences de la vie SPÉCIALITÉ : Biologie de la conservation Sujet de la thèse : Caractérisation génétique du genre Iris évoluant dans la Méditerranée Orientale. Présentée par : Nour ABDEL SAMAD Pour obtenir le grade de DOCTEUR ÈS SCIENCES Soutenue le 21/09/2016 Devant le jury composé de : Dr. Georges TOHME Président Dr. Teresa GARNATJE Rapporteur Dr. Gianluigi BACCHETTA Rapporteur Dr. Sophie NADOT Examinateur Dr. Laure EL CHAMY Examinateur Dr. Sonja SILJAK-YAKOVLEV Directeur de thèse Dr. Magda BOU DAGHER KHARRAT Directeur de thèse Titre : Caractérisation Génétique du Genre Iris évoluant dans la Méditerranée Orientale. Mots clés : Iris, Oncocyclus, région Est-Méditerranéenne, relations phylogénétiques, status taxonomique. Résumé : Le genre Iris appartient à la famille des L’approche scientifique est basée sur de nombreux Iridacées, il comprend plus de 280 espèces distribuées outils moléculaires et génétiques tels que : l’analyse de à travers l’hémisphère Nord. -

Starch Research Page URL: Last Updated: 14-April-2004 / Du Ethnobotanical Leaflets Starch Research Page
Journal Contents Back Issues Book Reviews Research Notes Careers Meetings Botany Resources Why study starch? The long and short of it is that size, shape, and optical and chemical properties of starch are useful in the determination of taxonomic species. Starch characteristics can be used in place of, or in combination with other features of the plant to facilitate the identification of macro archaeological plant remains, or even small pieces of modern plant materials which for one reason or the other are lacking the traditional "key" taxonomic characters. As in the case of pollen and phytolith analyses, the study of fossil starch grains can help the ethnobotanist to better understand the diet of ancient human cultures. The taxonomic usefulness of starch was long ago noted by Edward Tyson Reichert (1913), whose monumental work on "The Differentiation and Specificity of Starches in Relation to Genera, Species, Etc." has today become a classic in its field. He writes: "It must have been recognized by Leeuwenhoek, and by many of the investigators of the earliest part of the last century, that starches from different sources are not morphologically identical, but if so it does not seem to have attracted any particular attention until the investigations of Fritzsche (Ann. d. Physik. u. Chernie, 1834, xxxii, 129). Fritzsche described the starches obtained from a variety of plants. He noted not only that the starches from different sources were different, but also that often the form was so characteristic as to determine the plant, or, at least, indicate the genus and family from which the specimen was obtained. -

Patterns in Evolution in Characters That Define Iris Subgenera And
Aliso: A Journal of Systematic and Evolutionary Botany Volume 22 | Issue 1 Article 34 2006 Patterns in Evolution in Characters That Define rI is Subgenera and Sections Carol A. Wilson Rancho Santa Ana Botanic Garden Follow this and additional works at: http://scholarship.claremont.edu/aliso Part of the Botany Commons Recommended Citation Wilson, Carol A. (2006) "Patterns in Evolution in Characters That Define rI is Subgenera and Sections," Aliso: A Journal of Systematic and Evolutionary Botany: Vol. 22: Iss. 1, Article 34. Available at: http://scholarship.claremont.edu/aliso/vol22/iss1/34 Aliso 22, pp. 425-433 © 2006, Rancho Santa Ana Botanic Garden PATTERNS OF EVOLUTION IN CHARACTERS THAT DEFINE IRIS SUBGENERA AND SECTIONS CAROL A. WILSON Rancho Santa Ana Botanic Garden, 1500 North College Avenue, Claremont, California 91711-3157, USA (carol. wilson@ cgu. edu) ABSTRACT Subgeneric groups have been circumscribed in Iris based on a small number of morphological characters. Recent DNA sequence data has indicated that several of the subgenera, sections, and series that have previously been delineated are paraphyletic or polyphyletic. The evolution of characters that have traditionally been used to distinguish sub generic and sectional groups within Iris was investigated by mapping these characters on a phylogenetic tree based on matK sequence data. Results indicate that rhizomes are pleisomorphic for the genus and that three bulb types have arisen independently. My analysis shows that sepal beards, sepal crests, and seed arils show extensive homoplasy. Most of the homoplasy seen is associated with the circumscription of polyphyletic subgeneric groups such as the beardless subgenus Limniris. Some additional homoplasy is due to diversity within supported clades or the historical use of a single character in circumscribing more than one subgeneric group. -

February 2019 Newsletter
February 2019 TAIS Newsletter Our 54th year Tucson Area Iris Society—established 1965 An Affiliate of the American Iris Society President’s Message It starts with the green. At long last the iris fans are showing themselves. My records say I should see some blooms in mid-March. I'm hoping for a great bloom season and looking forward to seeing what hybridizer Bob Van Liere has to share with us at our February 9th meeting. - Kevin Kartchner P. S. Have you seen the irises blooming in front of Mesquite Valley Growers? ‘Spice Trader’ (Painter 2010) (AM, HM) ...“If you wish to ensure an iris is available if Hummingbird Iris Garden, Prescott yours would be eaten by a groundhog, sharing is Photo by Sue Clark, 2018 a good idea. You may want a piece back Inside this issue: someday”… - from the AIS Wiki Minutes from the January 2 meeting, Bronze Iris photos Upcoming Events Photos from the January 3 meeting February 9 meeting - Iris breeder Bob Van Liere of Iris4U Gardens in Denver will speak at 1 PM. Doors open at noon. Members are asked to bring a finger Summary of shade cloth, 4-6 fertilizing, and tips food to share and to help set up at noon. Murphy-Wilmot Branch Library, 530 N Wilmot Road, large room. All are welcome. Treasurer’s Report 7 March 9 meeting - Sue Clark presenting a visual tour of Longwood Gardens at 1 PM. Murphy-Wilmot Branch Library, 530 N Wilmot Road, small room. Bronze, Brass, and Gold 7 irises article April 13 - TAIS Iris Show, Murphy-Wilmot Library, 9 AM.